La evolución biológica es el proceso continuo de transformación de las especies a través de cambios producidos en sucesivas generaciones, y que se ve reflejado en el cambio de las frecuencias alélicas de una población.
Charles Darwin, padre de la teoría de la evolución por selección natural
Generalmente se denomina evolución a cualquier proceso de cambio en el tiempo. En el contexto de las Ciencias de la vida, la evolución es un cambio en el perfil genético de una población de individuos, que puede llevar a la aparición de nuevas especies, a la adaptación a distintos ambientes o a la aparición de novedades evolutivas.
A menudo existe cierta confusión entre hecho evolutivo y teoría de la evolución. Se denomina hecho evolutivo al hecho científico de que los seres vivos están emparentados entre sí y han ido transformándose a lo largo del tiempo. La teoría de la evolución es el modelo científico que describe la transformación evolutiva y explica sus causas.
Charles Darwin y Alfred Russel Wallace propusieron la selección natural como principal mecanismo de la evolución. Actualmente, la teoría de la evolución combina las propuestas de Darwin y Wallace con las leyes de Mendel y otros avances genéticos posteriores; por eso es llamada Síntesis Moderna o Teoría Sintética. En el seno de esta teoría, la evolución se define como un cambio en la frecuencia de los alelos en una población a lo largo de las generaciones. Este cambio puede ser causado por una cantidad de mecanismos diferentes: selección natural, deriva genética, mutación, migración (flujo genético). La Teoría Sintética recibe una aceptación general en la comunidad científica, aunque también ciertas críticas. Ha sido enriquecida desde su formulación, en torno a 1940, por avances en otras disciplinas relacionadas, como la biología molecular, la genética del desarrollo o la paleontología.
El Lamarckismo, la suposición de que el fenotipo de un organismo puede dirigir de alguna forma el cambio del genotipo en sus descendientes, es una posición teórica ya indefendible, en la medida en que es positivamente incompatible con lo que sabemos sobre la herencia; y también porque todos los intentos por hallar pruebas de observación o experimentales, han fracasado.
El creacionismo, la posición de que en un grado u otro, los seres vivos tienen un autor personal consciente (léase Dios), es una posición religiosa o filosófica que no puede probarse científicamente, y no es por tanto una teoría científica. No obstante, en el marco de la cultura popular protestante y anglosajona, algunos se esfuerzan por presentarlo como tal; pero la comunidad científica en su conjunto considera tales intentos como una forma de propaganda religiosa.
Teoría científica
La evolución biológica es un fenómeno natural real, observable y comprobable empíricamente. La llamada Síntesis Evolutiva Moderna es una robusta teoría que actualmente proporciona explicaciones y modelos matemáticos sobre los mecanismos generales de la evolución o los fenómenos evolutivos, como la adaptación o la especiación. Como cualquier teoría científica, sus hipótesis están sujetas a constante crítica y comprobación experimental.
Dobzhansky, uno de los fundadores de la Síntesis moderna, definió la evolución del siguiente modo: "La evolución es un cambio en la composición genética de las poblaciones. El estudio de los mecanismos evolutivos corresponde a la genética poblacional." [1] .
La síntesis moderna de la evolución se basa en tres aspectos fundamentales:
La ascendencia común de todos los organismos de un único ancestro.
El origen de nuevos caracteres en un linaje evolutivo.
Los mecanismos por los que algunos caracteres persisten mientras que otros desaparecen.
Origen y desarrollo temprano de la vida
El origen de la vida
Artículo principal: Origen de la vida
El origen de la vida, aunque atañe al estudio de los seres vivos, es un tema que realmente no es explicado en la teoría de la síntesis moderna de la evolución; pues ésta última sólo se ocupa del cambio en los seres vivos, y no de la creación y los cambios (evolución a moléculas más complejas) e interacciones de las moléculas orgánicas de las que procede.
No se sabe mucho sobre las etapas más tempranas y previas al desarrollo de la vida, y los intentos realizados para tratar de desvelar la historia más temprana del origen de la vida, generalmente se enfocan en el comportamiento de las macromoléculas, particularmente el ARN, y el comportamiento de sistemas complejos.
Sin embargo, si se esta de acuerdo que todos los organismos existentes comparten ciertas características, incluyendo la estructura celular y el código genético; los que estarían relacionados con el origen de la vida. (Para los científicos que consideran a los virus como seres vivos, si bien los mismos no tienen una estructura celular, evolucionaron a partir de organismos que sí las poseían, probablemente comportándose originalmente como transposones).
Ascendencia común
Artículo principal: Ascendencia común
A partir de estas semejanzas, los científicos interpretan que ellas indican y serían la evidencia de que todos los seres vivos existentes comparten un "ancestro común", el cual ya había desarrollado los procesos celulares más fundamentales; aunque no hay acuerdo en la comunidad científica sobre la relación específica de los tres dominios de la vida (Archaea, Bacteria, Eukaryota). Siendo desde la teoría del ancestro común, el comienzo de las explicaciones que son dadas por la teoría de la síntesis moderna de la evolución; en relación a la historia evolutiva de la vida.
Así, a pesar de que los orígenes de la vida nos son todavía desconocidos en su totalidad, otros hitos relacionados a la historia evolutiva de la vida sí son bien sabidos. La aparición de la fotosíntesis oxigénica (hace alrededor de 3000 millones de años) y el posterior surgimiento de una atmósfera rica en oxígeno y no reductora, puede rastrearse a través de depósitos laminares de hierro, y bandas rojas posteriores producto de los óxidos de hierro. Éste fue un requisito necesario para el desarrollo de la respiración celular aeróbica, la cual se cree que emergió hace aproximadamente 2000 millones de años. En los últimos mil millones de años, organismos pluricelulares simples, tanto plantas como animales, comenzaron a aparecer en los océanos. Poco después del surgimiento de los primeros animales, la explosión Cámbrica (un período breve de diversificación animal sin paralelo y notable, documentado en los fósiles encontrados en los sedimentos en Burgess Shale) vio la creación de la mayoría de los bauplans, o plan tipo, de los animales modernos. Hace alrededor de 500 millones de años, las plantas y hongos colonizaron la tierra, y fueron seguidos rápidamente por los artrópodos y otros animales, llevando al desarrollo de los ecosistemas terrestres con los que estamos familiarizados.
El surgimiento de nuevos caracteres y variación
Mecanismos de la herencia
En la época de Darwin, los científicos no estaban de acuerdo sobre cómo se heredan las características. Actualmente, el origen de la mayoría de las características hereditarias puede ser trazado hasta entidades persistentes llamadas genes, codificados en moléculas lineales llamadas ADN. El ADN varía entre los miembros de una misma especie y también sufre cambios o mutaciones, o variaciones producidas a través de procesos como la Recombinación genética.
Mutación
Artículo principal: Mutación
Darwin no conocía la fuente de las variaciones en los organismos individuales, pero observó que parecían ocurrir aleatoriamente. En trabajos posteriores se atribuyó la mayor parte de estas variaciones a la mutación. La mutación es un cambio permanente y transmisible en material genético (usualmente el ADN o el ARN) de una célula, que puede ser producida por errores de copia en el material genético durante la división celular y por la exposición a radiación, químicos o virus, o puede ocurrir deliberadamente bajo el control celular durante procesos como la meiosis o la hipermutación. En los organismos multicelulares, las mutaciones pueden dividirse en mutaciones germinales, que se transmiten a la descendencia, y las mutaciones somáticas, que (cuando son accidentales) generalmente conducen a malformaciones o muerte de células y pueden producir cáncer.
¿Por qué son importantes las mutaciones?
Las mutaciones introducen nuevas variaciones genéticas, siendo la principal fuente de evolución. En la teoría sintética, la mutación tiene el papel de generar diversidad genética sobre la cual actúa la selección natural, y también la deriva. Las mutaciones que afectan a la eficacia biológica del portador, y por tanto son objeto de la selección natural, pueden ser deletéreas (negativas) o beneficiosas. Las mutaciones beneficiosas son las menos frecuentes, aunque se conocen muchos ejemplos que afectan a rasgos variadísimos, como la resistencia a enfermedades o a estrés, la longevidad, el tamaño, la capacidad para metabolizar nuevas sustancias, una cicatrización eficiente de las heridas, etc. La mayor parte de las mutaciones son mutaciones neutras; no afectan las oportunidades de supervivencia y reproducción de los organismos, y se acumulan con el tiempo a una velocidad más o menos constante.
La mayoría de los biólogos creen que la adaptación ocurre fundamentalmente por etapas, mediante la acumulación por selección natural de variaciones genéticas ventajosas de efecto relativamente pequeño. Las macromutaciones, por el contrario, producen efectos drásticos, fuera del rango de variación normal de la especie. Se ha propuesto que quizá hayan sido responsables de ciertos rasgos adaptativos o de la aparición de novedades evolutivas, aunque, dado que las mutaciones suelen tener efectos muy nocivos o letales, esta vía se considera actualmente poco frecuente.
Recombinación genética
Artículo principal: Recombinación genética
La recombinación genética es el proceso mediante el cual la información genética se redistribuye, con la cual se produce variación en la descendencia y diversidad dentro de cada especie.
Variaciones en la expresión de los genes, involucrados en la herencia
También existen formas de variación hereditaria que no están basadas en cambios de la información genética. El proceso que produce estas variaciones deja intacta la información genética y es con frecuencia reversible. Este proceso es llamado herencia epigenética que resulta de la trasmisión de secuencias de información no-ADN a través de la meiosis o mitosis; y puede incluir fenómenos como la metilación del ADN o la herencia estructural. Se sigue investigando si estos mecanismos permiten la producción de variaciones específicas beneficiosas en respuesta a señales ambientales. De ser éste el caso, algunas instancias de la evolución podrían ocurrir fuera del cuadro típicamente darwiniano, que evitaría cualquier conexión entre las señales ambientales y la producción de variaciones hereditarias; aunque recordando que indirectamente el origen del proceso en si mismo estarían involucrados genes, como por ejemplo los genes de la enzima ADN-metiltransferasa, histonas, etc.
Sobrevivencia diferenciada de características
Al mismo tiempo que la mutación puede crear nuevos alelos, otros factores influencian la frecuencia de los alelos existentes. Estos factores hacen que algunas características se hagan frecuentes mientras que otras disminuyen o se pierden completamente. De los procesos conocidos que influyen en la persistencia de una característica, o más precisamente, en la frecuencia de un alelo podemos mencionar:
Selección natural
Deriva genética
Flujo genético
Selección natural
Artículo principal: Selección natural
La selección natural consiste en la reproducción diferencial de los individuos, según su dotación genética, y generalmente como resultado del ambiente. Existe selección natural cuando hay diferencias en eficacia biológica entre los individuos de una población, es decir, cuando su contribución en descendientes es desigual. La eficacia biológica puede desglosarse en componentes como la supervivencia (la mortalidad diferencial es la tasa de sobrevivencia de individuos hasta la edad de reproducción), la fertilidad, la fecundidad, etc.
La selección natural puede dividirse en dos categorías:
La sexual ocurre cuando los organismos más atractivos para el sexo opuesto debido a sus características se reproducen más y aumentan la frecuencia de estas características en el patrimonio genético común.
La ecológica ocurre en el resto de las circunstancias (habilidad para obtener o procesar alimento, capacidad de ocultación, huída o de defensa, capacidad para resistir fluctuaciones ambientales, etc.)
La selección natural trabaja con mutaciones en diferentes formas:
La purificadora o de fondo elimina las mutaciones perniciosas de una población.
La positiva aumenta la frecuencia de mutaciones benéficas.
La de balanceo mantiene las variaciones dentro de una población a través de mecanismos tales como:
La sobredominancia o vigor híbrido,
La selección dependiente de la frecuencia,
El papel central de la selección natural en la teoría de la evolución ha dado origen a una fuerte conexión entre ese campo y el estudio de la ecología.
Las mutaciones que no se ven afectadas por la selección natural son llamadas mutaciones neutras. Su frecuencia en la población está dictada por su tasa de mutación, por la deriva genética y el flujo genético. Se entiende que la secuencia de ADN de un organismo, en ausencia de selección, sufre una acumulación estable de mutaciones neutras. El efecto probable de mutación es la propuesta de que un gen que no está bajo selección será destruido por las mutaciones acumuladas. Éste es un aspecto de la llamada degradación genómica.
La selección de organismos por sus características deseables, cuando es provocada por el hombre, por ejemplo para la agricultura es llamada selección artificial.
La evolución baldwiniana se refiere a la forma en que los seres vivos capaces de adaptarse durante su vida, pueden producir nuevas fuerzas de selección.
Deriva genética
Artículo principal: Deriva genética
La deriva genética describe las fluctuaciones aleatorias en la frecuencia de los alelos. Esto es de especial importancia en poblaciones reducidas, donde las posibilidades de fluctuación de una generación a la siguiente son grandes. Estas fluctuaciones en la frecuencia de los alelos entre generaciones sucesivas puede producir la desaparición de algunos alelos de una población. Dos poblaciones separadas que parten de la misma frecuencia de alelos pueden derivar por fluctuación aleatoria en dos poblaciones divergentes con diferente conjunto de alelos (por ejemplo, alelos presentes en una población y que desaparecieron en la otra).
Muchos aspectos de la deriva genética dependen del tamaño de la población (generalmente abreviada como N). En las poblaciones reducidas, la deriva genética puede producir grandes cambios en la frecuencia de alelos de una generación a la siguiente, mientras que en las grandes poblaciones, los cambios en la frecuencia de los alelos son generalmente muy pequeños. La importancia relativa de la selección natural y la deriva genética en la determinación de la suerte de las nuevas mutaciones también depende del tamaño de la población y de la presión por la selección: Cuando N × s (tamaño de la población multiplicado por la presión por la selección) es pequeña, predomina la deriva genética. Así, la selección natural es más eficiente en grandes poblaciones o dicho de otra forma, la deriva genética es más poderosa en las poblaciones reducidas. Finalmente, el tiempo que le toma a un alelo fijarse en una población por deriva genética (es decir, el tiempo que toma el que todos los individuos de la población tengan ese alelo) depende del tamaño de la población: mientras más pequeña la población, menos tiempo toma la fijación del alelo.
Los efectos de la deriva genética son pequeños en la mayoría de las poblaciones naturales, pero pueden revestir especial importancia cuando tiene lugar la formación de una población a partir de muy pocos individuos o efecto fundador, o cuando las poblaciones quedan reducidas a muy pocos individuos, es decir, pasan a través de un cuello de botella.
Efecto fundador: Es un proceso frecuente en algunas islas oceánicas, que son colonizadas por unos pocos individuos que genéticamente son poco representativos con respecto a la población de la que derivan.
Un ejemplo que ilustra este efecto fundador se encuentra en el grupo religioso amish, fundado en 1771 en Pensilvania por unos pocos matrimonios. En la actualidad el 13% de las 17000 personas que forman el grupo portan en su genotipo un alelo que en homocigosis provoca enanismo y polidactilia. El número de casos registrados en esta población corresponde prácticamente a la totalidad de casos detectados en toda la población mundial. Se piensa que estas 17000 personas descienden de muy pocos individuos, algunos de los cuales eran portadores de este alelo.
Cuello de botella:Se produce cuando una situación en la que, debido a condiciones ambientales adversas u otras circunstancias, la población se reduce drásticamente. Con posterioridad recupera su número, pero a partir de un corto número de individuos. Esta situación puede implicar la desaparición de determinados alelos aleatoriamente o que aumente la frecuencia de otros que en la anterior situación estaban menos representados.
Microevolución y macroevolución
Artículos principales: Microevolución y Macroevolución
Microevolución es un término usado para referirse a cambios de las frecuencias génicas en pequeña escala, en una población durante el transcurso de varias generaciones. Estos cambios pueden deberse a un cierto número de procesos: mutación, flujo génico, deriva génica, así como también por selección natural. La genética de poblaciones es la rama de la biología que provee la estructura matemática para el estudio de los procesos de la microevolución, como el color de la piel en la población Mundial.
Los cambios a mayor escala, desde la especiación (aparición de una nueva especie) hasta las grandes transformaciones evolutivas ocurridas en largos períodos de tiempo, son comúnmente denominados Macroevolución (por ejemplo, los anfibios que evolucionaron a partir de un grupo de peces óseos). Los biólogos no acostumbran hacer una separación absoluta entre macroevolución y microevolución, pues consideran que macroevolución es simplemente microevolución acumulada y sometida a un rango mayor de circunstancias ambientales. Una minoría de teóricos, sin embargo, considera que los mecanismos de la teoría sintética para la microevolución no bastan para hacer esa extrapolación y que se necesitan otros mecanismos. La teoría de los equilibrios puntuados, propuesta por Gould y Eldredge, intenta explicar ciertas tendencias macroevolutivas que se observan en el registro fósil.
Especiación y extinción
Artículos principales: Especiación y Extinción
La especiación es la aparición de una o más especies a partir de una pre-existente. Existen varios mecanismos por los cuales esto puede ocurrir. La especiación alopátrica comienza cuando una subpoblación de una especie queda aislada geográficamente, por ejemplo por fragmentación del hábitat o migración. La especiación simpátrica ocurre cuando una especie nueva emerge en la misma región geográfica. La especiación peripátrica, propuesta por Ernst Mayr, es un tipo de especiación que existe entre los extremos de la especiación alopátrica y simpátrica. La especiación peripátrica es un soporte fundamental de la teoría del Equilibrio puntuado. La especiación parapátrica donde las especies ocupan áreas biogográficas aledañas pero hay un flujo genético bajo.
La extinción es la desaparición de las especies. El momento de la extinción es considerado generalmente como la muerte del último individuo perteneciente a una especie. La extinción no es un proceso inusual medido en tiempo geológico - las especies son creadas por la especiación y desaparecen a través de la extinción.
Biología evolutiva
Artículo principal: Biología evolutiva
La Biología evolutiva es un subcampo de la biología que se ocupa de la ascendencia común y evolución biológica de las especies, así como de sus cambios en el tiempo. La biología evolucionista es una especie de meta campo debido a que incluye científicos de muchas disciplinas tradicionales con orientación a la taxonomía. Por ejemplo, generalmente incluye científicos especializados en organismos particulares tales como la ornitología y la utiliza como medio para responder a preguntas generales sobre la evolución.
La biología evolutiva es una disciplina académica independiente que surgió en los años 1930 y 40 como resultado de la síntesis evolutiva moderna. Sin embargo, es en los años 1970 y 80 que un importante número de universidades crearon departamentos de biología evolutiva.
Historia del pensamiento evolucionista
Artículo principal: Historia del pensamiento evolucionista
La idea de una evolución biológica ha existido desde épocas remotas, notablemente entre los Helénicos como Epicuro, pero la teoría moderna no se estableció hasta llegados los siglos XVIII y XIX, con la contribución de científicos como Christian Pander, Jean-Baptiste Lamarck y Charles Darwin. En el siglo XVIII la oposición entre fijismo y transformismo es ambigua. Algunos autores, por ejemplo, admiten la transformación de las especies limitada a los géneros, pero niegan la posibilidad de pasar de un género a otro. Otros naturalistas hablan de "progresión" en la naturaleza orgánica, pero es muy difícil determinar si con ello hacen referencia a una transformación real de las especies o se trata, simplemente, de una modulación de la clásica idea de la scala naturae. Lamarck es el primero en formular explícitamente una teoría de la evolución, pero no fue hasta la publicación del El Origen de las Especies de Charles Darwin cuando el hecho de la evolución comenzó a ser ampliamente aceptado. Una carta de Alfred Russel Wallace, en la cual revelaba su propio descubrimiento de la selección natural, impulsó a Darwin a publicar su trabajo en evolución. Por lo tanto, a veces se comparte el crédito con Wallace por la teoría de la evolución (a veces llamada Teoría de Darwin-Wallace).
A pesar de que la teoría de Darwin pudo sacudir profundamente la opinión científica con respecto al desarrollo de la vida (e incluso resultando en una pequeña revolución social), no pudo explicar la fuente de variación existente entre las especies, y la propuesta de Darwin de la existencia de un mecanismo hereditario (pangénesis) no satisfizo a la mayoría de los biólogos. No fue recién hasta fines del siglo XIX y comienzos del XX, que estos mecanismos pudieron establecerse.
Cuando se "redescubrió" alrededor del 1900 el trabajo de Gregor Mendel sobre la naturaleza de la herencia que databa de fines del siglo XIX, se estableció una discusión entre los Mendelianos (Charles Benedict Davenport) y los biométricos Walter Frank Raphael Weldon y Karl Pearson), quienes insistían en que la mayoría de los caminos importantes para la evolución debían mostrar una variación continua que no era explicable a través del análisis mendeliano. Finalmente, los dos modelos fueron conciliados y fusionados, principalmente a través del trabajo del biólogo y estadístico R.A. Fisher. Este enfoque combinado, que empleaba un modelo estadístico riguroso a las teorías de Mendel de la herencia vía genes, se dio a conocer en los años 1930 y 1940 y se conoce como la teoría sintética de la evolución.
En los años de la década de 1940, siguiendo el experimento de Griffith, Avery, McCleod y McCarty lograron identificar de forma definitiva al ácido desoxirribonucléico (ADN) como el "principio transformante" responsable de la transmisión de la información genética. En 1953, Francis Crick y James Watson publicaron su famoso trabajo sobre la estructura del ADN, basado en la investigación de Rosalind Franklin y Maurice Wilkins. Estos desarrollos iniciaron la era de la biología molecular y transformaron el entendimiento de la evolución en un proceso molecular: la mutación de segmentos de ADN (ver evolución molecular).
A mediados de la década de 1970, Motoo Kimura formuló la teoría neutralista de la evolución molecular, estableciendo de manera firme la importancia de la deriva génica como el mayor mecanismo de la evolución. Hasta la fecha continúan los debates en esta área de investigación. Uno de los debates más importantes es sobre la teoría del equilibrio puntuado, una teoría propuesta por Niles Eldredge y Stephen Jay Gould para explicar la escasez de formas transicionales entre especies.
Evolución y sistemas éticos y sociales
La teoría de la evolución por acción de la selección natural también ha sido adoptada como fundamento para varios sistemas éticos y sociales, como el Darwinismo social, el cual mantiene que la supervivencia del más apto explica y justifica las diferencias de bienestar y éxito entre las sociedades, las personas y la eugenesia, que claman que la civilización humana estaba revirtiendo la selección natural permitiendo que los menos aptos sobrevivieran y se procrearan en exceso con respecto a los más aptos. Después de que las atrocidades del Holocausto fueran vinculadas con la eugenesia, la opinión pública científica dejó de ver de manera favorable la relación entre la selección natural y el Darwinismo social y la eugenesia (a pesar de que tampoco había sido realmente aceptada universalmente en el pasado).
Algunos creacionistas, como Kent Hovind, creen que la evolución es la base para el Nazismo, Comunismo, Marxismo, la alabanza a la Madre Tierra, racismo, etc.
La noción de que los humanos comparten ancestros comunes con otros animales, también afectó la manera en la que algunas personas ven la relación entre los humanos y otras especies. Muchos de los defensores de los derechos humanos mantienen que si los animales y humanos son de la misma naturaleza, por lo que entonces los derechos no pueden ser distintos para los humanos.
Evolución y religión
Artículos principales: Creacionismo, Diseño inteligente, y Evolución teísta
Antes de que la geología se convirtiera en una ciencia, a principios del siglo XIX, tanto las religiones occidentales como los científico descontaban o condenaban de manera dogmática y casi unánime cualquier propuesta que implicara que la vida es el resultado de un proceso evolutivo. Sin embargo, a medida que la evidencia geológica empezó a acumularse en todo el mundo, un grupo de científicos comenzó a cuestionar si una interpretación literal de la creación relatada en la Biblia Judeo-Cristiana podía reconciliarse con sus descubrimientos (y sus implicaciones). Algunos geólogos religiosos, como Dean William Auckland en Inglaterra, Edward Hitchcock en Estados Unidos y Hugo Millar en Escocia siguieron justificando la evidencia geológica y fósil solo en términos de un Diluvio universal; pero una vez que Charles Darwin publicara su Origen de las Especies en 1859 la opinión científica comenzó a alejarse rápidamente de la interpretación literal de la Biblia.
Este debate temprano acerca de la validez literal de la Biblia no se llevó a cabo tras puertas cerradas, y desestabilizó la opinión educativa en ambos continentes. Eventualmente, instigó una contrarreforma que tomó la forma de un renacimiento religioso en ambos continentes entre 1857 y 1860.
A pesar que la teoría de la evolución ha sido demostrada científicamente, algunos grupos, principalmente en Estados Unidos, interpretan en las Escrituras que solo un ser supremo pudo crear directamente a los humanos y a otros animales como especies separadas y acabadas. Este punto de vista es comúnmente llamado creacionismo, y sigue siendo defendido por algunos grupos religiosos, particularmente los protestantes estadounidenses; principalmente a través de una forma de creacionismo contemporáneo llamado Diseño inteligente. Esto ha llevado a un duro conflicto entre la creación y evolución en la historia de la educación pública de este país, aunque actualmente más bien es un fenómeno local en algunos estados; ya que es obligatoria la enseñanza de la teoría de evolución. (aunque cabe destacar que también a afectado a otros países, por ejemplo, en el año 2005 en Italia hubo un intento de suspensión de la enseñanza de la teoría de la evolución).
En respuesta a la aceptación científica de la teoría de la evolución, muchos religiosos y filósofos han tratado de unificar los puntos de vista científico y religioso, ya sea de manera formal o informal; a través de un "creacionismo pro-evolución". Así por ejemplo algunos religiosos han adoptado un enfoque creacionista desde la evolución teísta o el creacionismo evolutivo, en donde Dios provee una chispa divina que inicia el proceso de la evolución, y (o) donde Dios creó el curso de la evolución.
Por ejemplo, a partir de 1950 la Iglesia Católica Romana tomó una posición neutral con respecto a la evolución con la encíclica Humani generis del Papa Pío XII. "El Magisterio de la Iglesia no prohíbe el que —según el estado actual de las ciencias y la teología— en las investigaciones y disputas, entre los hombres más competentes de entrambos campos, sea objeto de estudio la doctrina del evolucionismo, en cuanto busca el origen del cuerpo humano en una materia viva preexistente —pero la fe católica manda defender que las almas son creadas inmediatamente por Dios—. ". El Papa Benedicto XVI ha afirmado que "existen muchas pruebas científicas en favor de la evolución, que se presenta como una realidad que debemos ver y que enriquece nuestro conocimiento de la vida y del ser como tal. Pero la doctrina de la evolución no responde a todos los interrogantes y sobre todo no responde al gran interrogante filosófico: ¿de dónde viene todo esto y cómo todo toma un camino que desemboca finalmente en el hombre?"[2] .
En los países o regiones en los cuales de la mayoría de la población mantiene fuertes creencias religiosas, el creacionismo posee un atractivo mucho mayor que en los países donde la mayoría de la gente posee creencias seculares. Desde los años 1920 hasta el presente en los Estados Unidos, han ocurrido varios ataques religiosos a la enseñanza de la teoría evolutiva, particularmente por parte de los cristianos fundamentalistas y protestantes; si bien entre los últimos esta no es una posición unánime.
miércoles, 28 de noviembre de 2007
GENETICA BASICA
¿QUE ES LA GENETICA?
La genética es la ciencia que estudia la variación y la transmisión de rasgos o características de una generación a la otra. En esta definición, la palabra variación se refiere a variación genética; esto significa, el rango de posibles valores para un rasgo cuando es influenciado por la herencia. La herencia es la transmisión de rasgos de los padres a la descendencia vía el material genético. Esta transmisión toma lugar en el momento de la fertilización en la reproducción, cuando un espermatozoide de toro se une con el óvulo de la vaca para producir un ternero con una composición genética única. Solamente mellizos idénticos poseen la composición genética idéntica debido a que ellos descienden de un solo óvulo fertilizado que ha sido separado en dos embriones durante la primera fase del desarrollo.¿QUE ES MEDIO AMBIENTE?El medio ambiente es generalmente entendido como los alrededores físicos del animal, luz, temperatura, ventilación y otros parámetros que pueden contribuir al confort físico del animal. Aún así, en genética, la palabra medio ambiente posee un significado más general. El medio ambiente es la combinación de todos los factores, con excepción de los genéticos, que pueden afectar la expresión de los genes. Por ejemplo, la producción de leche de la vaca se encuentra afectada por la edad al parto, la época del parto, la nutrición y muchos otros factores. Por lo tanto, vacas que tengan una composición genética similar o igual producirán diferentes cantidades de leche cuando se encuentren sometidas a diferentes medios ambientes. Por ejemplo, el desempeño en la lactancia de un par de mellizas idénticas variará drásticamente si dos terneras son separadas al nacer y enviadas a distintos países. Aún así, puede haber una gran diferencia en producción de leche entre estas gemelas cuando se ubiquen en dos explotaciones lecheras separadas pero dentro de la misma área, cada una teniendo diferentes niveles de manejo.
TRANSMISION DEL MATERIAL GENETICO
Macho y hembraLos testículos del toro y los ovarios de la vaca producen las células reproductoras por una serie de divisiones celulares que separan el número de cromosomas en una célula. El espermatozoide y el ovario contienen solamente un miembro del par de cromosomas. Por lo tanto, las células de la vaca y del toro contienen 60 cromosomas (2n = 60), pero el espermatozoide en el semen y el óvulo en los ovarios contienen solamente 30 cromosomas (n=30, Figura 2). Los dos principios básicos de la transmisión de un rasgo (ejm. sexo) son los siguientes (Figura 2):1) Separación de los pares de cromosomas durante la formación de las células reproductoras;2) Unión del espermatozoide con el óvulo para crear una nueva célula con un conjunto único de cromosomas.Para 29 pares de cromosomas, ambos miembros son visualmente idénticos. De todas formas, para uno de los pares, un miembro es mucho más largo; es llamado cromosoma X, y el miembro más corto es llamado cromosoma Y. Todos los óvulos llevan el cromosoma X, pero el espermatozoide puede llevar ya sea el cromosoma X o el Y. Durante la división celular para formar las células reproductoras, cada miembro del par de cromosomas va hacia una célula por separado. Como resultado, 50% de los espermatozoides llevarán el cromosoma X y el otro 50%, el cromosoma Y. Si por casualidad un espermatozoide que lleva el cromosoma Y fertiliza un óvulo, la descendencia será macho. Si la descendencia recibe dos cromosomas X, se desarrollará una hembra (Figura 2). Es importante darse cuenta que es imposible predecir el sexo de la descendencia al momento del apareamiento (inseminación); aún así, podemos predecir que, en promedio, 50% de la descendencia serán machos y 50% hembras.Rasgos cualitativosLos rasgos cualitativos tienden a caer dentro de categorías discretas. Generalmente solo uno o unos pocos genes poseen un gran efecto sobre los rasgos cualitativos. El medio ambiente tene generalmente un pequeño papel al influenciar la categoría dentro de la que el animal cae. En este caso, el fenotipo de un animal refleja su genotipo. Ejemplos de rasgos cualitativos son:* Color de pelo;* Defectos hereditarios como enanismo;* Presencia o ausencia de cuernos;* Tipo sanguíneo.Rasgos cuantitativosLos rasgos cuantitativos difieren de los cualitativos de dos formas importantes:1) Se encuentran influenciados por muchos pares de genes;2) La expresión fenotípica es influenciada más fuertemente por el medio ambiente que en el caso de los rasgos cualitativos.Muchos de los rasgos de importancia económica importante en el ganado lechero son cuantitativos:* Producción de leche;* Composición de la leche;* Conformación (también llamado tipo);* Eficiencia de conversión de alimento;* Resistencia a enfermedades.La influencia combinada de muchos genes y el efecto del medio ambiente en los rasgos cuantitativos hacen que sea mucho más difícil el determinar el genotipo exacto que en el caso de la mayoría de los rasgos cualitativos. Algunas veces, el fenotipo del animal nos dice muy poco acerca de su genotipo. Por ejemplo, un registro de lactancia solamente dice una fracción de la información acerca de el mérito genético de la vaca para producción de leche.Qué hace que el genotipo de una vaca sea único?Cuando se forman los óvulos, ellos reciben uno de los dos miembros del par de cromosomas. Por lo tanto, un cromosoma en particular en un óvulo puede ser el primer o el segundo miembro del par de cromosomas de los padres. Existen solamente dos tipos de óvulos para un gen en particular. Si en lugar de un par de cromosomas, consideramos dos, cuál es el número de diferentes óvulos?. En otras palabras, cuál es el número total de combinaciones cromosómicas posibles?. La situación es la misma que la de arrojar dos monedas al mismo tiempo. El número de posibles combinaciones es: dos posibles valores para la primera moneda multiplicado por los dos posibles valores de la segunda = 2 x 2 = 22 = 4 diferentes posibilidades. El número de diferentes genotipos para un óvulo es cuatro y la probabilidad de una combinación en particular de cromosomas es de 1/4. Esto es también verdad para el número de posibles genotipos en las células reproductoras masculinas. Por lo tanto, cuando uno de cuatro posibles clases de espermatozoides fertiliza uno de cuatro posibles combinaciones de óvulos el número de descendientes genéticamente diferentes es 4 x 4 = 16 (ejm., 22 x 22 ). Por lo tanto, las chances de que un genotipo en particular se presente en el recién nacido es 1/16.Cuando los 30 pares de cromosomas del ganado lechero se separan durante la formación de las células reproductoras y luego se vuelven a unir en el momento de la fertilización, el número total de posibles combinaciones cromosómicas es 230 x 230 = 1.152.900.000.000.000.000, cada uno siendo único. Con este número de posibilidades para cada apareamiento, es fácil entender por qué dos individuos no son iguales en una población, aún cuando tengan el mismo padre.
¿QUE ES LA GENETICA?
La genética es la ciencia que estudia la variación y la transmisión de rasgos o características de una generación a la otra. En esta definición, la palabra variación se refiere a variación genética; esto significa, el rango de posibles valores para un rasgo cuando es influenciado por la herencia. La herencia es la transmisión de rasgos de los padres a la descendencia vía el material genético. Esta transmisión toma lugar en el momento de la fertilización en la reproducción, cuando un espermatozoide de toro se une con el óvulo de la vaca para producir un ternero con una composición genética única. Solamente mellizos idénticos poseen la composición genética idéntica debido a que ellos descienden de un solo óvulo fertilizado que ha sido separado en dos embriones durante la primera fase del desarrollo.
¿QUE ES MEDIO AMBIENTE?
El medio ambiente es generalmente entendido como los alrededores físicos del animal, luz, temperatura, ventilación y otros parámetros que pueden contribuir al confort físico del animal. Aún así, en genética, la palabra medio ambiente posee un significado más general. El medio ambiente es la combinación de todos los factores, con excepción de los genéticos, que pueden afectar la expresión de los genes. Por ejemplo, la producción de leche de la vaca se encuentra afectada por la edad al parto, la época del parto, la nutrición y muchos otros factores. Por lo tanto, vacas que tengan una composición genética similar o igual producirán diferentes cantidades de leche cuando se encuentren sometidas a diferentes medios ambientes. Por ejemplo, el desempeño en la lactancia de un par de mellizas idénticas variará drásticamente si dos terneras son separadas al nacer y enviadas a distintos países. Aún así, puede haber una gran diferencia en producción de leche entre estas gemelas cuando se ubiquen en dos explotaciones lecheras separadas pero dentro de la misma área, cada una teniendo diferentes niveles de manejo.
GENOTIPO Y FENOTIPO
El genotipo de un animal representa el gen o grupo de genes responsable por un rasgo en particular. En un sentido más general, el genotipo describe todo el grupo de genes que un individuo ha heredado.Como contraste, el fenotipo es el valor que toma un rasgo; en otras palabras, es lo que puede ser observado o medido. Por ejemplo, el fenotipo puede ser la producción individual de leche de una vaca, el porcentaje de grasa en la leche o el puntaje de clasificación por conformación.Existe una diferencia importante entre genotipo y fenotipo. El genotipo es esencialmente una característica fija del organismo; permanece constante a lo largo de la vida del animal y no es modificado por el medio ambiente. Cuando solamente uno o un par de genes son responsables por un rasgo, el genotipo permanece generalmente sin cambios a lo largo de la vida del animal (ejm. color de pelo). En este caso, el fenotipo otorga una buena indicación de la composición genética del individuo. Aún así, para algunos rasgos, el fenotipo cambia constantemente a lo largo de la vida del individuo como respuesta a factores ambientales. En este caso, el fenotipo no es un indicador confiable del genotipo. Esto generalmente se presenta cuando muchos genes se encuentran involucrados en la expresión de un rasgo tal como producción de leche. Como resultado, la producción de leche de una vaca depende de:Producción fenotípica de leche = G + E, donde:G es el mérito genético de la vaca para producción de leche (el efecto de los genes);E se refiere al efecto del manejo de la vaca y medio ambiente.EL MATERIAL GENETICO El material genético se encuentra localizado en el núcleo de cada célula del cuerpo. A excepción de las células reproductoras (espermatozoides y óvulos) y algunas otras excepciones (glóbulos rojos sanguíneos), las células contienen dos copias del material genético completo del animal. Cuando la célula se divide, el material genético se organiza en una serie de estructuras largas en forma de fibras llamadas cromosomas (Figura 1). En las células del cuerpo, cada cromosoma posee una contraparte que posee el mismo largo y forma (con la excepción de los cromosomas que determinan el sexo) y contienen la información genética del mismo rasgo. Estos dos cromosomas son miembros de un par de cromosomas, uno derivado del padre y otro de la madre. El número de pares de cromosomas es típico de una especie y es generalmente abreviado con la letra "n". Por ejemplo, en humanos n=23, en cerdos n=19, en vacas n=30. Por lo tanto las células en el cuerpo humano, cerdos y vacas contienen 2n=46, 38 y 60 cromosomas, respectivamente.Los genes se encuentran localizados a lo largo de los cromosomas. Un gen es la unidad funcional básica de la herencia; esto significa que contiene la información genética que es responsable por la expresión de un rasgo en particular. El largo completo de un cromosoma puede dividirse en miles de estas unidades funcionales, cada una responsable de un rasgo en particular.Un gen se compone de ácido desoxiribonucleico o ADN. La función del ADN es la de llevar la información necesaria para la síntesis de proteínas. A medida que las proteínas son sintetizadas y que el ADN se replica a sí mismo, el número de células del cuerpo se incrementa (crecimiento) y las células pueden especializarse dentro de diferentes funciones específicas (desarrollo) en las que algunos genes se expresan otros no. Por ejemplo, las células de la piel (tejido especializado) contienen todo el material genético necesario para recrear un individuo, pero los únicos genes especializados que se expresan en estas células son los responsables por la formación y el color del pelo.
TRANSMISION DEL MATERIAL GENETICO
Macho y hembraLos testículos del toro y los ovarios de la vaca producen las células reproductoras por una serie de divisiones celulares que separan el número de cromosomas en una célula. El espermatozoide y el ovario contienen solamente un miembro del par de cromosomas. Por lo tanto, las células de la vaca y del toro contienen 60 cromosomas (2n = 60), pero el espermatozoide en el semen y el óvulo en los ovarios contienen solamente 30 cromosomas (n=30, Figura 2). Los dos principios básicos de la transmisión de un rasgo (ejm. sexo) son los siguientes (Figura 2):1) Separación de los pares de cromosomas durante la formación de las células reproductoras;2) Unión del espermatozoide con el óvulo para crear una nueva célula con un conjunto único de cromosomas.Para 29 pares de cromosomas, ambos miembros son visualmente idénticos. De todas formas, para uno de los pares, un miembro es mucho más largo; es llamado cromosoma X, y el miembro más corto es llamado cromosoma Y. Todos los óvulos llevan el cromosoma X, pero el espermatozoide puede llevar ya sea el cromosoma X o el Y. Durante la división celular para formar las células reproductoras, cada miembro del par de cromosomas va hacia una célula por separado. Como resultado, 50% de los espermatozoides llevarán el cromosoma X y el otro 50%, el cromosoma Y. Si por casualidad un espermatozoide que lleva el cromosoma Y fertiliza un óvulo, la descendencia será macho. Si la descendencia recibe dos cromosomas X, se desarrollará una hembra (Figura 2). Es importante darse cuenta que es imposible predecir el sexo de la descendencia al momento del apareamiento (inseminación); aún así, podemos predecir que, en promedio, 50% de la descendencia serán machos y 50% hembras.Rasgos cualitativosLos rasgos cualitativos tienden a caer dentro de categorías discretas. Generalmente solo uno o unos pocos genes poseen un gran efecto sobre los rasgos cualitativos. El medio ambiente tene generalmente un pequeño papel al influenciar la categoría dentro de la que el animal cae. En este caso, el fenotipo de un animal refleja su genotipo. Ejemplos de rasgos cualitativos son:* Color de pelo;* Defectos hereditarios como enanismo;* Presencia o ausencia de cuernos;* Tipo sanguíneo.5.3 Rasgos cuantitativosLos rasgos cuantitativos difieren de los cualitativos de dos formas importantes:1) Se encuentran influenciados por muchos pares de genes;2) La expresión fenotípica es influenciada más fuertemente por el medio ambiente que en el caso de los rasgos cualitativos.Muchos de los rasgos de importancia económica importante en el ganado lechero son cuantitativos:* Producción de leche;* Composición de la leche;* Conformación (también llamado tipo);* Eficiencia de conversión de alimento;* Resistencia a enfermedades.La influencia combinada de muchos genes y el efecto del medio ambiente en los rasgos cuantitativos hacen que sea mucho más difícil el determinar el genotipo exacto que en el caso de la mayoría de los rasgos cualitativos. Algunas veces, el fenotipo del animal nos dice muy poco acerca de su genotipo. Por ejemplo, un registro de lactancia solamente dice una fracción de la información acerca de el mérito genético de la vaca para producción de leche.Qué hace que el genotipo de una vaca sea único?Cuando se forman los óvulos, ellos reciben uno de los dos miembros del par de cromosomas. Por lo tanto, un cromosoma en particular en un óvulo puede ser el primer o el segundo miembro del par de cromosomas de los padres. Existen solamente dos tipos de óvulos para un gen en particular. Si en lugar de un par de cromosomas, consideramos dos, cuál es el número de diferentes óvulos?. En otras palabras, cuál es el número total de combinaciones cromosómicas posibles?. La situación es la misma que la de arrojar dos monedas al mismo tiempo. El número de posibles combinaciones es: dos posibles valores para la primera moneda multiplicado por los dos posibles valores de la segunda = 2 x 2 = 22 = 4 diferentes posibilidades. El número de diferentes genotipos para un óvulo es cuatro y la probabilidad de una combinación en particular de cromosomas es de 1/4. Esto es también verdad para el número de posibles genotipos en las células reproductoras masculinas. Por lo tanto, cuando uno de cuatro posibles clases de espermatozoides fertiliza uno de cuatro posibles combinaciones de óvulos el número de descendientes genéticamente diferentes es 4 x 4 = 16 (ejm., 22 x 22 ). Por lo tanto, las chances de que un genotipo en particular se presente en el recién nacido es 1/16.Cuando los 30 pares de cromosomas del ganado lechero se separan durante la formación de las células reproductoras y luego se vuelven a unir en el momento de la fertilización, el número total de posibles combinaciones cromosómicas es 230 x 230 = 1.152.900.000.000.000.000, cada uno siendo único. Con este número de posibilidades para cada apareamiento, es fácil entender por qué dos individuos no son iguales en una población, aún cuando tengan el mismo padre.La Herencia, perspectiva histórica Durante gran parte de la historia de la humanidad las personas desconocían los detalles científicos de la concepción y de como trabajaba la herencia. Por cierto los niños eran concebidos y por cierto se veía que existía una semejanza entre padres e hijos, pero los mecanismos no eran conocidos. Los filósofos griegos tenían varias ideas: Teofrasto (371-287 a.C.) comprendía la diferencia entre las flores masculinas y femeninas, decía que "los machos debían ser llevados a las hembras" dado que los machos "hacían madurar y persistir" a las flores hembras; Hipócrates (460?- 377? a.C.) especuló, que las "semillas" se producían en diferentes partes del cuerpo y se transmitían a los hijos al momento de la concepción, y Aristóteles pensó que el semen masculino y el semen femenino (así se llamaba al flujo menstrual) se mezclaban en la concepción, algunos pensaban que ni siquiera este tipo de mezclas eran necesarias, las formas "simples" (gusano, moscas...) nacían por generación espontánea.Durante los 1700s, Anton van Leeuwenhoek (1632-1723, para los no holandeses lii-uen-huuk seria una pronunciación bastante aceptable; sus aportes y los de otros pioneros pueden leerse en una magnífica novelización) descubre "animálculos" en el esperma humano y de otros animales. Algunos de los que miraban por los primeros microscopios soñaron ver un "pequeño hombrecito" (homúnculo) dentro de cada espermatozoide. Sostuvieron que la única contribución de la hembra para la próxima generación era proveer el ambiente para su desarrollo. En oposición la escuela de los ovistas creía que el futuro hombre estaba en el óvulo, y que el espermatozoide solo lo estimulaba, creían también que había huevos para hembras y para machos.La pangénesis sostenía la idea que machos y hembras forman "pangenes" en cada órgano. Estos "pangenes" se movían a través de la sangre a los genitales y luego a los recién nacidos. El concepto, originado en los griegos influenció a la biología hasta hace solo unos 100 años. Los términos "sangre azul", "consanguíneo", "hermano de sangre", "mezcla de sangre", "sangre gitana" y otros similares surgen de estos conceptos. Francis Galton, un primo de Charles Darwin, desecho experimentalmente la pangénesis.Las teoría de la mezcla ("Blending theories") suplantó a la de los espermistas y ovistas durante el siglo 19. La mezcla de óvulos y espermatozoides daban como resultado la progenie que era una "mezcla" ("blend") de las características de los padres. Las células sexuales se conocían colectivamente como gametos. De acuerdo con la teoría de la mezcla, cuando un animal de color negro se cruzaba con uno blanco la progenie debía ser gris y, a menudo, este no era el resultado. La teoría de la mezcla obviaba, entre otras, explicar el salto de generación de algunas características.Charles Darwin en su teoría de la evolución, se vio forzado a reconocer que la mezcla no era un factor (o al menos no el factor principal) y sugirió que la ciencia, en la mitad de los 1800s, no tenía la respuesta correcta al problema. La respuesta vino de un contemporáneo, Gregor Mendel, si bien Darwin nunca conoció el trabajo de Mendel.Conceptos básicos de la genéticaResulta útil recordar algunos conceptos previos para comprender los experimentos de Mendel, aunque este monje no haya tenido conocimiento de los genes o los cromosomas... Meiosis: división celular que origina 4 células con la mitad de la dotación cromosómica de la célula orginal (haploides). Los cromosomas homólogos se separan y cada célula (gameta) recibe uno de los homólogos del par.Carácter: característica observable y transmitida por los genes, ejemplo: color de las floresFenotipo : propiedades observables del genotipo y en el cual contribuye el medio ambiente.Cromosomas Homólogos: cromosomas que se aparean durante la meiosis. Poseen igual longitud, posición del centrómero y comparten los mismos genes. Excepción : cromosomas X e Y que no comparten las características anteriores pero sí se consideran homólogos por aparearse en la meiosis.Gen (del griego genos= nacimiento) son segmentos específicos de ADN (cromosoma) responsable de un determinado carácter; son la unidad funcional de la herencia. El botánico danés Wilhelm Johannsen (1857 - 1927) acuño este nombre, en 1909, para nombrar a los elemente de Mendel (también acuñó "fenotipo", "genotipo" y "selección").Alelo: Formas alternativas de un gen en un mismo locus. Por ejemplo 2 posibles alelos en el locus v de la cebada son v y V. El término de alelo ó alelomorfo fue acuñado por William Bateson; literalmente significa "forma alternativa".Locus: es el lugar específico de un gen en un cromosoma.Homocigoto: organismo que tiene dos copias o alelos iguales de un gen en los dos homólogos, también llamado raza pura.Heterocigoto: cuando los dos alelos son diferentes, en este caso el alelo dominante es el que se expresa.MENDELMendel razonó que un organismo apto para los experimentos genéticos debería tener :una serie de características diferentes, fácilmente estudiables y con dos o tres fenotipos diferentes.la planta debía autofertilizarse y tener una estructura floral que limite los contactos accidentales, de crecimiento rápido y con gran número de descendientes.Los descendientes de las plantas autofertilizadas debían ser fértiles.El organismo experimental de Mendel fue la arveja común (Pisum sativum, familia Leguminosae), que tiene una flor que normalmente se autopoliniza. La parte masculina de la flor se llama antera, produce el polen, que contiene los gametos masculinos. La parte femenina de la flor es el Gineceo, formado por estigma, estilo, y el ovario. El óvulo (gameto femenino) es producido en el ovario. El proceso de polinización (la transferencia de polen de la antera al estigma) ocurre, en el caso de la arveja, antes de la apertura de la flor. Del grano de polen crece un tubo (tubo polínico) que permite al núcleo viajar a través del estigma y el estilo, y eventualmente llegar al ovario. Las paredes del ovario formarán las futuras vainas (fruto: legumbre) y los óvulos fecundados las semillas. Ver ciclo animado de plantas.Muchas flores permiten la polinización cruzada, lo cual puede dificultar los estudios si se desconoce las características de la planta masculina. Dado que las flores de las arvejas el estigma y las anteras están completamente encerrados y, a diferencia de la mayoría de las flores no se abren hasta ser fecundadas, es decir luego de la autopolinización, la genética de los progenitores puede ser comprendida mas fácilmente. Los embriones autofecundados de las arvejas desarrollan sin dificultad.Para los entrecruzamientos Mendel abrió el pimpollo antes de la maduración y retiró las anteras con pinzas evitando la autopolinización. Luego las polinizó artificialmente, espolvoreando el estigma con polen recogido de otras plantas.Mendel probó las 34 variedades de arvejas disponibles a través de los vendedores de semillas. Mendel buscó caracteres con rasgos bien definidos y alternativos constantes, que constituyeran razas puras. Las arvejas de jardín fueron plantadas y estudiadas durante ocho años a fin de comprobar que el rasgo observado se mantenía constante a lo largo de varias generaciones. Así, Mendel aisló 7 pares de caracteres que eran razas puras: cada carácter estudiado se presentaba en dos variantes, tales como: altura de la planta (alta o baja), superficie de la semilla (lisa o rugosa), forma de la vaina (inflada o contraída), forma de la vaina y otras (ver esquema a continuación). En sus experimentos Mendel uso unas 28.000 plantas de arvejas.La contribución de Mendel fue excepcional en razón del enfoque metodológico utilizado para definir el problema, el uso de variables claramente entendibles y la aplicación de las matemática (estadística) al resultado experimental. Usando plantas de arvejas y el método estadístico, Mendel fue capaz de demostrar que los caracteres pasan de los padres a los hijos a través de la herencia de los genes.El principio de la segregación Mendel primero estudió la herencia de la forma de la semilla. Un cruzamiento relacionado a un solo carácter se denomina monohibridación. Mendel cruzó una raza pura de plantas con semillas lisas con una raza pura de otra que siempre producía semillas rugosas (60 fertilizaciones en 15 plantas). Todas las semillas resultantes resultaron lisas.Al año siguiente, Mendel plantó esas semillas y permitió que las mismas se autofecunden. Recogió 7324 semillas en total: 5474 lisas y 1850 rugosas. Para sistematizar el registro de datos, las generaciones fueron nombradas y numeradas. La generación parental se denomina como P. Los descendientes de la generación P son la generación F1 (la primera filial). La autofecundación de la generación de F1 produce la generación F2 (la segunda filial).P1:lisa X rugosaF1 :todas lisasF2 :5474 lisas y 1850 rugosasLo mismo sucedió con cada par de caracteres elegidos: cuando cepas puras de plantas con semillas amarillas se cruzan con razas puras de plantas con semillas verdes, todos los descendientes fueron plantas con semillas amarillas. Los padres del entrecruzamiento son la generación P1, y los descendientes representan la generación F1. Cuando los miembros de la generación F1 se entrecruzaron, Mendel recobro muchos descendientes amarillas, y algunos verdes. Luego del análisis estadístico de la generación F2, Mendel determinó que la relación entre plantas con semillas amarillas/verdes era 3:1. Las plantas con semillas verdes no aparecían en la primera generación F1, y se encontraban en la segunda F2 y sucesivas generaciones.Cruzamiento monohíbrido entre semillas amarillas (dominantes) y verdes (recesivo). Vea este Mendel concluyó que el carácter estudiado estaba gobernado por factores discretos (separables) y que el rasgo del carácter que aparece en la F1 es el dominante. Los factores se heredaban a pares, teniendo cada generación un par de los mismos. Actualmente nos referimos a esos factores como alelos. El hecho de que los caracteres se hereden de a pares permiten explicar el fenómeno observado del "salto" de una generación.Los caracteres dominantes fueron definidos por Mendel como aquellos que aparecen en la primera generación( F1) en los entrecruzamientos entre dos especies puras. Las letras mayúsculas se usan generalmente como notación para los caracteres dominantesLos caracteres recesivos son los que "saltan" una generación, y se observan únicamente cuando el carácter dominante esta ausente. Las letras minúsculas se usan generalmente como notación para los caracteres recesivos.Las plantas de Mendel exhibían dominancia completa, en las cuales las expresiones fenotípicas de los alelos eran dominantes o recesivas, sin "caracteres intermedios".La Meiosis, un proceso desconocido en los días de Mendel, explica como se heredan los caracteres:Esquema de la meiosis y formación de los gametos. Imagen modificada deSumario de los resultados de MendelLos descendientes F1 muestran solo uno de los caracteres de los padres, y siempre el mismo carácter.El carácter que no se observa en F1 reaparece en F2 en aproximadamente un 25% de los descendientes.El carácter no cambia cuando pasa a la descendencia: no se mezclan en ningún descendiente y se comportan como unidades separadas.Los cruzamientos recíprocos demostraron que cada progenitor contribuye de manera igual a la descendencia.El término fenotipo se refiere al conjunto de caracteres que se expresan o sea a la apariencia externa, mientras que el término genotipo se refiere a la totalidad genética del individuo .Machos y hembras contribuyen equitativamente a la formación del material genético de la descendencia: por lo tanto el numero de factores que determinan un carácter es probablemente dos (la solución mas simple).El Principio de la Segregación o Primera Ley de Mendel, propone la separación de los factores apareados durante la formación de los gametos, donde cada gameto recibe uno u otro factor durante su formación. Los organismos portan dos factores (alelos) por cada carácter. Estos factores se separan durante la formación de los gametos.Una versión en hipertexto (en Alemán o Ingles) del trabajo original de Mendel en 1865 se consigue siguiendo este enlace.Consecuencias de la segregaciónAlelos: se sabe ahora que cualquier gen presenta dos formas diferentes o alelos.Homo- y Heterocigosis: determinada por la combinación de los dos alelos de un gen.Fenotipo: expresión de las características genéticas o genotipo.Cuadro de PUNNETEs un mecanismo muy útil a la hora de considerar las posibles combinaciones de gametos. Por ejemplo, en la F1 todas las plantas del cruzamiento monohíbrido entre plantas altas y bajas dieron altas. El cuadro de Punnett permite calcular el resultado de la F2:Cruzamiento de pruebaPara probar la hipótesis de que los alelos están en pares y se separan en la formación de gametas se llevó a cabo un experimento adicional: se cruzó la F1 (semillas lisas) con la raza pura paterna de semillas rugosas (padre homocigota recesivo) a lo que se denominó
CRUZAMIENTO DE PRUEBA.
En un cruzamiento de prueba se cruzan un genotipo desconocido que muestra el carácter dominante con el padre homocigota recesivo. Lo que se pretende demostrar es si el genotipo desconocido es homocigota dominante o heterocigota para ese carácter. Si se producen dos fenotipos distintos quiere decir que el progenitor desconocido era heterocigota para ese carácter. Si por el contrario aparece un solo fenotipo es homocigoto.Cruzamiento dihíbridoMendel entendió que era necesario realizar su experimento en una situación más compleja y realizó experimentos siguiendo dos caracteres de las semillas: forma y color. Un entrecruzamiento concerniente a dos caracteres se conoce como cruzamiento dihíbrido en oposición al cruzamiento de una sola característica o, monohíbrido.La generación F2 resultante no muestra la característica relación fenotípica 3:1 dominante: recesivo. Los dos caracteres, si consideramos que se heredan independientemente, "calzan" dentro del principio de la segregación. En vez de los 4 posibles genotipos de un monohíbrido, el cruzamiento dihíbrido tiene 16 posibles genotipos.Cruzamientos con dos caracteresLas semillas lisas (S) son dominantes respecto a la semillas rugosos (s).El color amarillo (Y) es dominante sobre el verde (y).Una vez más, la meiosis nos ayuda a entender el comportamiento de los alelos.Métodos, Resultados y ConclusionesEl gráfico superior es de Genetics pages en McGill UniversityMendel partió de razas puras que tenían plantas con semillas lisas y amarillas, y las cruzó con razas puras de plantas con semillas verdes y arrugadas. Todas las semillas de la generación F1 tenían semillas lisas y amarillas. Las plantas de la generación F2 se obtuvieron por autofertilización, y produjeron cuatro fenotipos:315 lisas y amarillas108 lisas verdes101 arrugadas amarillas32 arrugadas verdesMendel analizó cada carácter por separado como si fuera que el otro carácter no estuviera presente. la relación 3:1 se veía separadamente y estaba de acuerdo con el Principio de Segregación. La segregación de los alelos S y s debían haber ocurrido independientemente de la separación de los alelos Y e y .La probabilidad de que un gameto tenga Y es 1/2; la probabilidad de cualquier gameto de tener S es 1/2. La probabilidad de que un gameto contenga ambos Y y S se calcula por el producto de las probabilidades individuales (o 1/2 X 1/2 = 1/4).La probabilidad de que dos gametos formen cualquier mezcla de estos alelos en su genotipo 1/4 X 1/4 (recuerde el producto de las probabilidades individuales).Por lo tanto, existen 16 posibilidades y, el tablero de Punnett tiene 16 casillas. Dado que hay mas posibilidades de combinaciones que producen el fenotipo liso y amarillo (SSYY, SsYy, SsYY, y SSYy), este fenotipo es mas común en la F2.De los resultados de su segundo experimento, Mendel formuló el Principio de la distribución independiente esto es, cuando se forman los gametos, los alelos de un gen para una característica dada se separan (segregan) independientemente de un alelo para otra característica. Si los caracteres se separan independientemente uno de otros durante la formación de los gametos, puede entenderse el resultado de un entrecruzamiento dihíbrido.Desde los tiempo de Mendel, los científicos descubrieron el cromosoma y el ADN, y actualmente se interpreta el principio de la distribución independiente como alelos de genes en diferentes cromosomas que se heredan independientemente durante la formación de los gametos. Esto no era del conocimiento de Mendel.MutaciónDe Vries en 1902 trabajando sobre la "hierba del asno" describió en ella fenómenos de herencia mendeliana, sin embargo de tanto en tanto aparecía una característica que no estaba ni en los padres ni en los antecesores de las plantas, dedujo de ello que estas característica surgían por un cambio el factor que determinaba el carácter (gen) y que este cambio se transmitía a la progenie como cualquier otro carácter hereditario. A este cambio lo denominó mutación y a los organismos que la mostraban mutantes, los alelos salvajes son los que están presentes en la mayoría de los individuos y dan el fenotipo esperado.Ni las leyes de Mendel ni el concepto de mutación fueron conocidos por Darwin, pero resulta claro que la combinación de características de los padres da resultados sobre los cuales puede actuar el proceso evolutivo y que las mutaciones (si bien raras) son una fuente constante de variaciones que posibilitan la evolución.Los diversos alelos existen porque cualquier gen está sometido a mutaciones, que ocurren cuando un gen cambia a una nueva forma estable y hereditaria. Las mutaciones son procesos aleatorios. Los alelos mutantes y salvajes residen en el mismo locus y se heredan de acuerdo a la genética mendeliana.Las Leyes de Mendel.-Primera Ley de Mendel o Principio de Ley de la Dominancia.- Esta Ley menciona que para cada característica hereditaria existen genes dominantes y recesivos. Sin importar cual padre contribuye con el carácter dominante el híbrido o heterocigoto siempre tendrá fenotipo dominante.Segunda Ley de Mendel o Principio de la Segregación de Caracteres.Un carácter hereditario se transmite como una unidad que no se combina, se diluye o se pierde al pasar de una generación a otra, sólo se segrega o se separa.Tercera Ley de Mendel o de Distribución Independiente. Anuncia que un par de alelos se distribuye en forma independiente de otro par de alelos. Los caracteres se heredan de manera independiente unos de otros.Material hereditarioEl material hereditario es un componente de las células que da las características a éstas, además de darles una actividad específica. En las células eucariontes se ubica dentro del núcleo celular. Recibe este nombre a causa de que al dividirse una célula en dos, este material se duplica y cada célula hija "hereda" una copia de él. También se le conoce como material genético.Descubrimientos a través de la historiaA mediados del siglo XIX, Gregorio Mendel determinó que el material hereditario funciona como partículas que conservan su identidad en el paso de una célula a otra al interior de los gametos. A estas "partículas" las llamó factores hereditarios.Al comenzar el siglo XX, los estudios de Mendel estaban siendo reconocidos. Había argumentos a favor de la teoría de que los factores hereditarios se encuentran dentro de una partícula llamada cromosoma. Estos pensamientos estaban favorecidos por la semejanza entre las teorías de Mendel y el comportamiento que se estaba observando microscópicamente en los cromosomas.Las investigaciones continuaron y luego, llevaron a concluir que los factores hereditarios, ahora llamados genes, estaban hechos de ADN. Una prueba de lo anterior sería que en todas las células de la misma especie, la cantidad de ADN es constante. En cambio en los gametos existe la mitad de este elemento. Ya en la década de 1960 se conocía modelos de la estructura de ésta molécula, como también se sabía como se duplicaba y como determinaba la función de una célula. Se logró establecer que la secuencia de nucleótidos presentes en el ADN determina a la secuencia de aminoácidos en las proteínas, llegando a conocer el papel que desempeña el nucleótido ARN. Para que los ribosomas logren sintetizar las proteínas, el ADN se copia mediante la transcripción, en una molécula de ARN, también llamada ARN mensajero. Este sale del núcleo y llega a los ribosomas, que a través de la traducción, leen el patrón del ARN y efectúan la síntesis de proteínas.
MUTACION DE LOS GENES
Las mutaciones fueron descritas por primera vez en 1901 por uno de los redescubridores de Mendel, el botánico holandés Hugo De Vries.Si esto se produce en la secuencia de nucleótidos que codifica un polipéptido particular, éste puede presentar un aminoácido cambiado en la cadena polipeptídica. Esta modificación puede alterar seriamente las propiedades de la proteína resultante. Cuando se produce una mutación durante la formación de los gametos, ésta se transmitirá a las siguientes generaciones. La mayoría de las mutaciones genéticas son perjudiciales para el organismo que las porta. Una modificación aleatoria es más fácil que deteriore y que no mejore la función de un sistema complejo como el de una proteína. Por esta razón, en cualquier momento, el número de sujetos que portan un gen mutante determinado se debe a dos fuerzas opuestas: la tendencia a aumentar debido a la propagación de individuos mutantes nuevos en una población, y la tendencia a disminuir debido a que los individuos mutantes no sobreviven o se reproducen menos que sus semejantes. Varias actuaciones humanas recientes, como la exposición a los rayos X con fines médicos, los materiales radiactivos y las mutaciones producidas por compuestos químicos, son responsables de su aumento.Por lo general, las mutaciones son recesivas, sus efectos perjudiciales no se expresan a menos que dos de ellos coincidan para dar lugar a una situación homocigótica. Esto es más probable en la procreación consanguínea, en el apareamiento de organismos muy relacionados que pueden haber heredado el mismo gen mutante recesivo de un antecesor común. Por esta razón, las enfermedades hereditarias son más frecuentes entre los niños cuyos padres son primos que en el resto de la población.
Mutaciones génicas.
Son las verdaderas mutaciones, porque se produce un cambio en la estructura del ADN. A pesar de todos los sistemas destinados a prevenir y corregir los posibles errores, estos pueden serlas mutaciones puntuales, que son el cambio de una de las bases de un par en el ADN, un cambio en una base nitrogenada puede alterar la estructura completa de la proteína.Tirosina arginina metioninaAUG GCU UACTirosina glutamina metioninaAUG GUU UAC
De vez en cuando se produce alguno en la réplica, bien por colocarse una Citosina (C) en lugar de una Timina (T), o una Adenina (A) en lugar de una Guanina (G); o bien porque el mecanismo de replicación se salta algunas bases y aparece una "mella" en la copia. O se unen dos bases de Timina, formando un dímero. Aunque se trate de un cambio de un nucleótido por otro, supondrá una alteración en la secuencia de un gen, que se traduce posteriormente en una modificación de la secuencia de aminoácidos de una proteína. Al transcribirse la mutación, al menos un triplete del ARNm , se encuentra modificado y su traducción da lugar a que se incorpore un aminoácido distinto del normal en la cadena polipeptídica. Es un cambio que aunque la mayoría de las veces va a ser perjudicial, en contadas ocasiones puede provocar que mejore un gen y gracias a esta característica se sintetice una proteína distinta , que tenga propiedades distintas o participe en la formación de estructuras más eficaces. En estos casos raros, pero esenciales para la evolución de las especies , los individuos portadores de la mutación poseen ventajas adaptativas respecto a sus congéneres , por lo que el gen mutado es posible que con el tiempo, y gracias a la selección natural, sustituya al gen original en la mayoría de los individuos que componen la población
Mutaciones cromosómicas
La sustitución de un nucleótido por otro no es el único tipo posible de mutación. Algunas veces se puede ganar o perder por completo un nucleótido. Además, es posible que se produzcan modificaciones más obvias o graves, o que se altere la propia forma y el número de los cromosomas. Una parte del cromosoma se puede separar, invertir y después unirse de nuevo al cromosoma en el mismo lugar. A esto se le llama inversión. Si el fragmento separado se une a un cromosoma distinto, o a un fragmento diferente del cromosoma original, el fenómeno se denomina translocación. Algunas veces se pierde un fragmento de un cromosoma que forma parte de una pareja de cromosomas homólogos, y este fragmento es adquirido por el otro. Entonces, se dice que uno presenta una deleción o deficiencia (dependiendo si el fragmento que se pierde es intersticial o terminal, respectivamente) y el otro una duplicación. Por lo general, las deficiencias o deleciones son letales en la condición homocigótica, y con frecuencia las duplicaciones también lo son. Las inversiones y las translocaciones suelen ser más viables, aunque pueden asociarse con mutaciones en los genes cerca de los puntos donde los cromosomas se han roto. Es probable que la mayoría de estos reordenamientos cromosómicos sean la consecuencia de errores en el proceso de sobrecruzamiento.
Errores de dusyución
Este tipo de mutaciones afectan a la dotación cromosómica de un individuo, es decir, los individuos que las presentan tienen en sus células un número distinto de cromosomas al que es propio de su especie. No son mutaciones propiamente dichas, porque no hay cambio de material genético, sino una aberración, la cual suele ser el resultado de una separación anormal de los cromosomas durante la meiosis, con lo que podemos encontrarnos individuos triploides (3n), tetraploides (4n), etc. Cuando en la meiosis fracasa la separación de una pareja de cromosomas homólogos. Esto puede originar gametos —y, por lo tanto, cigotos— con cromosomas de más, y otros donde faltan uno o más cromosomas. En el hombre, existen varios síndromes provocados por la no separación de una pareja de cromosoma homólogos durante la meiosis, con lo cual permanecen unidos y se desplazan juntos a un mismo gameto provocando lo que se denomina trisomía, es decir un individuo con un cromosoma triplicado. Los individuos con un cromosoma de más se denominan trisómicos, y aquellos en los que falta uno, monosómicos. Ambas situaciones tienden a producir incapacidades graves. Por ejemplo, las personas con síndrome de Down son trisómicas, con tres copias del cromosoma 21.En la meiosis fracasa a veces la separación de un grupo completo de cromosomas; es decir, se origina un gameto con el doble del número normal de cromosomas. Si dicho gameto se une con otro que contiene el número normal de cromosomas, el descendiente tendrá tres grupos de cromosomas homólogos en lugar de los dos habituales. Si se unen dos gametos con el doble del número normal de cromosomas, el descendiente estará dotado de cuatro grupos homólogos. Los organismos con grupos adicionales de cromosomas reciben el nombre de poliploides. La poliploidía es el único proceso conocido por el cual pueden surgir especies nuevas en una generación única. Se han observado poliploides viables y fértiles casi exclusivamente en organismos hermafroditas, como la mayoría de las plantas con flores y algunos invertebrados Por lo general, las plantas poliploides son mayores y más robustas que sus antecesoras diploides Estos poliploides así formados son genéticamente muy interesantes en las plantas cultivadas, y hoy en día la mayoría de variedades gigantes de fresones, tomates, trigo, ... que existen en el mercado, tienen este origen.. Algunas veces se originan fetos poliploides en la raza humana, pero fallecen en una fase precoz del desarrollo fetal y se produce un aborto.En los siguientes esquemas, tenemos las trisomías más frecuentes tanto en los autosomas, como en los cromosomas sexuales.
La genética es la ciencia que estudia la variación y la transmisión de rasgos o características de una generación a la otra. En esta definición, la palabra variación se refiere a variación genética; esto significa, el rango de posibles valores para un rasgo cuando es influenciado por la herencia. La herencia es la transmisión de rasgos de los padres a la descendencia vía el material genético. Esta transmisión toma lugar en el momento de la fertilización en la reproducción, cuando un espermatozoide de toro se une con el óvulo de la vaca para producir un ternero con una composición genética única. Solamente mellizos idénticos poseen la composición genética idéntica debido a que ellos descienden de un solo óvulo fertilizado que ha sido separado en dos embriones durante la primera fase del desarrollo.¿QUE ES MEDIO AMBIENTE?El medio ambiente es generalmente entendido como los alrededores físicos del animal, luz, temperatura, ventilación y otros parámetros que pueden contribuir al confort físico del animal. Aún así, en genética, la palabra medio ambiente posee un significado más general. El medio ambiente es la combinación de todos los factores, con excepción de los genéticos, que pueden afectar la expresión de los genes. Por ejemplo, la producción de leche de la vaca se encuentra afectada por la edad al parto, la época del parto, la nutrición y muchos otros factores. Por lo tanto, vacas que tengan una composición genética similar o igual producirán diferentes cantidades de leche cuando se encuentren sometidas a diferentes medios ambientes. Por ejemplo, el desempeño en la lactancia de un par de mellizas idénticas variará drásticamente si dos terneras son separadas al nacer y enviadas a distintos países. Aún así, puede haber una gran diferencia en producción de leche entre estas gemelas cuando se ubiquen en dos explotaciones lecheras separadas pero dentro de la misma área, cada una teniendo diferentes niveles de manejo.
TRANSMISION DEL MATERIAL GENETICO
Macho y hembraLos testículos del toro y los ovarios de la vaca producen las células reproductoras por una serie de divisiones celulares que separan el número de cromosomas en una célula. El espermatozoide y el ovario contienen solamente un miembro del par de cromosomas. Por lo tanto, las células de la vaca y del toro contienen 60 cromosomas (2n = 60), pero el espermatozoide en el semen y el óvulo en los ovarios contienen solamente 30 cromosomas (n=30, Figura 2). Los dos principios básicos de la transmisión de un rasgo (ejm. sexo) son los siguientes (Figura 2):1) Separación de los pares de cromosomas durante la formación de las células reproductoras;2) Unión del espermatozoide con el óvulo para crear una nueva célula con un conjunto único de cromosomas.Para 29 pares de cromosomas, ambos miembros son visualmente idénticos. De todas formas, para uno de los pares, un miembro es mucho más largo; es llamado cromosoma X, y el miembro más corto es llamado cromosoma Y. Todos los óvulos llevan el cromosoma X, pero el espermatozoide puede llevar ya sea el cromosoma X o el Y. Durante la división celular para formar las células reproductoras, cada miembro del par de cromosomas va hacia una célula por separado. Como resultado, 50% de los espermatozoides llevarán el cromosoma X y el otro 50%, el cromosoma Y. Si por casualidad un espermatozoide que lleva el cromosoma Y fertiliza un óvulo, la descendencia será macho. Si la descendencia recibe dos cromosomas X, se desarrollará una hembra (Figura 2). Es importante darse cuenta que es imposible predecir el sexo de la descendencia al momento del apareamiento (inseminación); aún así, podemos predecir que, en promedio, 50% de la descendencia serán machos y 50% hembras.Rasgos cualitativosLos rasgos cualitativos tienden a caer dentro de categorías discretas. Generalmente solo uno o unos pocos genes poseen un gran efecto sobre los rasgos cualitativos. El medio ambiente tene generalmente un pequeño papel al influenciar la categoría dentro de la que el animal cae. En este caso, el fenotipo de un animal refleja su genotipo. Ejemplos de rasgos cualitativos son:* Color de pelo;* Defectos hereditarios como enanismo;* Presencia o ausencia de cuernos;* Tipo sanguíneo.Rasgos cuantitativosLos rasgos cuantitativos difieren de los cualitativos de dos formas importantes:1) Se encuentran influenciados por muchos pares de genes;2) La expresión fenotípica es influenciada más fuertemente por el medio ambiente que en el caso de los rasgos cualitativos.Muchos de los rasgos de importancia económica importante en el ganado lechero son cuantitativos:* Producción de leche;* Composición de la leche;* Conformación (también llamado tipo);* Eficiencia de conversión de alimento;* Resistencia a enfermedades.La influencia combinada de muchos genes y el efecto del medio ambiente en los rasgos cuantitativos hacen que sea mucho más difícil el determinar el genotipo exacto que en el caso de la mayoría de los rasgos cualitativos. Algunas veces, el fenotipo del animal nos dice muy poco acerca de su genotipo. Por ejemplo, un registro de lactancia solamente dice una fracción de la información acerca de el mérito genético de la vaca para producción de leche.Qué hace que el genotipo de una vaca sea único?Cuando se forman los óvulos, ellos reciben uno de los dos miembros del par de cromosomas. Por lo tanto, un cromosoma en particular en un óvulo puede ser el primer o el segundo miembro del par de cromosomas de los padres. Existen solamente dos tipos de óvulos para un gen en particular. Si en lugar de un par de cromosomas, consideramos dos, cuál es el número de diferentes óvulos?. En otras palabras, cuál es el número total de combinaciones cromosómicas posibles?. La situación es la misma que la de arrojar dos monedas al mismo tiempo. El número de posibles combinaciones es: dos posibles valores para la primera moneda multiplicado por los dos posibles valores de la segunda = 2 x 2 = 22 = 4 diferentes posibilidades. El número de diferentes genotipos para un óvulo es cuatro y la probabilidad de una combinación en particular de cromosomas es de 1/4. Esto es también verdad para el número de posibles genotipos en las células reproductoras masculinas. Por lo tanto, cuando uno de cuatro posibles clases de espermatozoides fertiliza uno de cuatro posibles combinaciones de óvulos el número de descendientes genéticamente diferentes es 4 x 4 = 16 (ejm., 22 x 22 ). Por lo tanto, las chances de que un genotipo en particular se presente en el recién nacido es 1/16.Cuando los 30 pares de cromosomas del ganado lechero se separan durante la formación de las células reproductoras y luego se vuelven a unir en el momento de la fertilización, el número total de posibles combinaciones cromosómicas es 230 x 230 = 1.152.900.000.000.000.000, cada uno siendo único. Con este número de posibilidades para cada apareamiento, es fácil entender por qué dos individuos no son iguales en una población, aún cuando tengan el mismo padre.
¿QUE ES LA GENETICA?
La genética es la ciencia que estudia la variación y la transmisión de rasgos o características de una generación a la otra. En esta definición, la palabra variación se refiere a variación genética; esto significa, el rango de posibles valores para un rasgo cuando es influenciado por la herencia. La herencia es la transmisión de rasgos de los padres a la descendencia vía el material genético. Esta transmisión toma lugar en el momento de la fertilización en la reproducción, cuando un espermatozoide de toro se une con el óvulo de la vaca para producir un ternero con una composición genética única. Solamente mellizos idénticos poseen la composición genética idéntica debido a que ellos descienden de un solo óvulo fertilizado que ha sido separado en dos embriones durante la primera fase del desarrollo.
¿QUE ES MEDIO AMBIENTE?
El medio ambiente es generalmente entendido como los alrededores físicos del animal, luz, temperatura, ventilación y otros parámetros que pueden contribuir al confort físico del animal. Aún así, en genética, la palabra medio ambiente posee un significado más general. El medio ambiente es la combinación de todos los factores, con excepción de los genéticos, que pueden afectar la expresión de los genes. Por ejemplo, la producción de leche de la vaca se encuentra afectada por la edad al parto, la época del parto, la nutrición y muchos otros factores. Por lo tanto, vacas que tengan una composición genética similar o igual producirán diferentes cantidades de leche cuando se encuentren sometidas a diferentes medios ambientes. Por ejemplo, el desempeño en la lactancia de un par de mellizas idénticas variará drásticamente si dos terneras son separadas al nacer y enviadas a distintos países. Aún así, puede haber una gran diferencia en producción de leche entre estas gemelas cuando se ubiquen en dos explotaciones lecheras separadas pero dentro de la misma área, cada una teniendo diferentes niveles de manejo.
GENOTIPO Y FENOTIPO
El genotipo de un animal representa el gen o grupo de genes responsable por un rasgo en particular. En un sentido más general, el genotipo describe todo el grupo de genes que un individuo ha heredado.Como contraste, el fenotipo es el valor que toma un rasgo; en otras palabras, es lo que puede ser observado o medido. Por ejemplo, el fenotipo puede ser la producción individual de leche de una vaca, el porcentaje de grasa en la leche o el puntaje de clasificación por conformación.Existe una diferencia importante entre genotipo y fenotipo. El genotipo es esencialmente una característica fija del organismo; permanece constante a lo largo de la vida del animal y no es modificado por el medio ambiente. Cuando solamente uno o un par de genes son responsables por un rasgo, el genotipo permanece generalmente sin cambios a lo largo de la vida del animal (ejm. color de pelo). En este caso, el fenotipo otorga una buena indicación de la composición genética del individuo. Aún así, para algunos rasgos, el fenotipo cambia constantemente a lo largo de la vida del individuo como respuesta a factores ambientales. En este caso, el fenotipo no es un indicador confiable del genotipo. Esto generalmente se presenta cuando muchos genes se encuentran involucrados en la expresión de un rasgo tal como producción de leche. Como resultado, la producción de leche de una vaca depende de:Producción fenotípica de leche = G + E, donde:G es el mérito genético de la vaca para producción de leche (el efecto de los genes);E se refiere al efecto del manejo de la vaca y medio ambiente.EL MATERIAL GENETICO El material genético se encuentra localizado en el núcleo de cada célula del cuerpo. A excepción de las células reproductoras (espermatozoides y óvulos) y algunas otras excepciones (glóbulos rojos sanguíneos), las células contienen dos copias del material genético completo del animal. Cuando la célula se divide, el material genético se organiza en una serie de estructuras largas en forma de fibras llamadas cromosomas (Figura 1). En las células del cuerpo, cada cromosoma posee una contraparte que posee el mismo largo y forma (con la excepción de los cromosomas que determinan el sexo) y contienen la información genética del mismo rasgo. Estos dos cromosomas son miembros de un par de cromosomas, uno derivado del padre y otro de la madre. El número de pares de cromosomas es típico de una especie y es generalmente abreviado con la letra "n". Por ejemplo, en humanos n=23, en cerdos n=19, en vacas n=30. Por lo tanto las células en el cuerpo humano, cerdos y vacas contienen 2n=46, 38 y 60 cromosomas, respectivamente.Los genes se encuentran localizados a lo largo de los cromosomas. Un gen es la unidad funcional básica de la herencia; esto significa que contiene la información genética que es responsable por la expresión de un rasgo en particular. El largo completo de un cromosoma puede dividirse en miles de estas unidades funcionales, cada una responsable de un rasgo en particular.Un gen se compone de ácido desoxiribonucleico o ADN. La función del ADN es la de llevar la información necesaria para la síntesis de proteínas. A medida que las proteínas son sintetizadas y que el ADN se replica a sí mismo, el número de células del cuerpo se incrementa (crecimiento) y las células pueden especializarse dentro de diferentes funciones específicas (desarrollo) en las que algunos genes se expresan otros no. Por ejemplo, las células de la piel (tejido especializado) contienen todo el material genético necesario para recrear un individuo, pero los únicos genes especializados que se expresan en estas células son los responsables por la formación y el color del pelo.
TRANSMISION DEL MATERIAL GENETICO
Macho y hembraLos testículos del toro y los ovarios de la vaca producen las células reproductoras por una serie de divisiones celulares que separan el número de cromosomas en una célula. El espermatozoide y el ovario contienen solamente un miembro del par de cromosomas. Por lo tanto, las células de la vaca y del toro contienen 60 cromosomas (2n = 60), pero el espermatozoide en el semen y el óvulo en los ovarios contienen solamente 30 cromosomas (n=30, Figura 2). Los dos principios básicos de la transmisión de un rasgo (ejm. sexo) son los siguientes (Figura 2):1) Separación de los pares de cromosomas durante la formación de las células reproductoras;2) Unión del espermatozoide con el óvulo para crear una nueva célula con un conjunto único de cromosomas.Para 29 pares de cromosomas, ambos miembros son visualmente idénticos. De todas formas, para uno de los pares, un miembro es mucho más largo; es llamado cromosoma X, y el miembro más corto es llamado cromosoma Y. Todos los óvulos llevan el cromosoma X, pero el espermatozoide puede llevar ya sea el cromosoma X o el Y. Durante la división celular para formar las células reproductoras, cada miembro del par de cromosomas va hacia una célula por separado. Como resultado, 50% de los espermatozoides llevarán el cromosoma X y el otro 50%, el cromosoma Y. Si por casualidad un espermatozoide que lleva el cromosoma Y fertiliza un óvulo, la descendencia será macho. Si la descendencia recibe dos cromosomas X, se desarrollará una hembra (Figura 2). Es importante darse cuenta que es imposible predecir el sexo de la descendencia al momento del apareamiento (inseminación); aún así, podemos predecir que, en promedio, 50% de la descendencia serán machos y 50% hembras.Rasgos cualitativosLos rasgos cualitativos tienden a caer dentro de categorías discretas. Generalmente solo uno o unos pocos genes poseen un gran efecto sobre los rasgos cualitativos. El medio ambiente tene generalmente un pequeño papel al influenciar la categoría dentro de la que el animal cae. En este caso, el fenotipo de un animal refleja su genotipo. Ejemplos de rasgos cualitativos son:* Color de pelo;* Defectos hereditarios como enanismo;* Presencia o ausencia de cuernos;* Tipo sanguíneo.5.3 Rasgos cuantitativosLos rasgos cuantitativos difieren de los cualitativos de dos formas importantes:1) Se encuentran influenciados por muchos pares de genes;2) La expresión fenotípica es influenciada más fuertemente por el medio ambiente que en el caso de los rasgos cualitativos.Muchos de los rasgos de importancia económica importante en el ganado lechero son cuantitativos:* Producción de leche;* Composición de la leche;* Conformación (también llamado tipo);* Eficiencia de conversión de alimento;* Resistencia a enfermedades.La influencia combinada de muchos genes y el efecto del medio ambiente en los rasgos cuantitativos hacen que sea mucho más difícil el determinar el genotipo exacto que en el caso de la mayoría de los rasgos cualitativos. Algunas veces, el fenotipo del animal nos dice muy poco acerca de su genotipo. Por ejemplo, un registro de lactancia solamente dice una fracción de la información acerca de el mérito genético de la vaca para producción de leche.Qué hace que el genotipo de una vaca sea único?Cuando se forman los óvulos, ellos reciben uno de los dos miembros del par de cromosomas. Por lo tanto, un cromosoma en particular en un óvulo puede ser el primer o el segundo miembro del par de cromosomas de los padres. Existen solamente dos tipos de óvulos para un gen en particular. Si en lugar de un par de cromosomas, consideramos dos, cuál es el número de diferentes óvulos?. En otras palabras, cuál es el número total de combinaciones cromosómicas posibles?. La situación es la misma que la de arrojar dos monedas al mismo tiempo. El número de posibles combinaciones es: dos posibles valores para la primera moneda multiplicado por los dos posibles valores de la segunda = 2 x 2 = 22 = 4 diferentes posibilidades. El número de diferentes genotipos para un óvulo es cuatro y la probabilidad de una combinación en particular de cromosomas es de 1/4. Esto es también verdad para el número de posibles genotipos en las células reproductoras masculinas. Por lo tanto, cuando uno de cuatro posibles clases de espermatozoides fertiliza uno de cuatro posibles combinaciones de óvulos el número de descendientes genéticamente diferentes es 4 x 4 = 16 (ejm., 22 x 22 ). Por lo tanto, las chances de que un genotipo en particular se presente en el recién nacido es 1/16.Cuando los 30 pares de cromosomas del ganado lechero se separan durante la formación de las células reproductoras y luego se vuelven a unir en el momento de la fertilización, el número total de posibles combinaciones cromosómicas es 230 x 230 = 1.152.900.000.000.000.000, cada uno siendo único. Con este número de posibilidades para cada apareamiento, es fácil entender por qué dos individuos no son iguales en una población, aún cuando tengan el mismo padre.La Herencia, perspectiva histórica Durante gran parte de la historia de la humanidad las personas desconocían los detalles científicos de la concepción y de como trabajaba la herencia. Por cierto los niños eran concebidos y por cierto se veía que existía una semejanza entre padres e hijos, pero los mecanismos no eran conocidos. Los filósofos griegos tenían varias ideas: Teofrasto (371-287 a.C.) comprendía la diferencia entre las flores masculinas y femeninas, decía que "los machos debían ser llevados a las hembras" dado que los machos "hacían madurar y persistir" a las flores hembras; Hipócrates (460?- 377? a.C.) especuló, que las "semillas" se producían en diferentes partes del cuerpo y se transmitían a los hijos al momento de la concepción, y Aristóteles pensó que el semen masculino y el semen femenino (así se llamaba al flujo menstrual) se mezclaban en la concepción, algunos pensaban que ni siquiera este tipo de mezclas eran necesarias, las formas "simples" (gusano, moscas...) nacían por generación espontánea.Durante los 1700s, Anton van Leeuwenhoek (1632-1723, para los no holandeses lii-uen-huuk seria una pronunciación bastante aceptable; sus aportes y los de otros pioneros pueden leerse en una magnífica novelización) descubre "animálculos" en el esperma humano y de otros animales. Algunos de los que miraban por los primeros microscopios soñaron ver un "pequeño hombrecito" (homúnculo) dentro de cada espermatozoide. Sostuvieron que la única contribución de la hembra para la próxima generación era proveer el ambiente para su desarrollo. En oposición la escuela de los ovistas creía que el futuro hombre estaba en el óvulo, y que el espermatozoide solo lo estimulaba, creían también que había huevos para hembras y para machos.La pangénesis sostenía la idea que machos y hembras forman "pangenes" en cada órgano. Estos "pangenes" se movían a través de la sangre a los genitales y luego a los recién nacidos. El concepto, originado en los griegos influenció a la biología hasta hace solo unos 100 años. Los términos "sangre azul", "consanguíneo", "hermano de sangre", "mezcla de sangre", "sangre gitana" y otros similares surgen de estos conceptos. Francis Galton, un primo de Charles Darwin, desecho experimentalmente la pangénesis.Las teoría de la mezcla ("Blending theories") suplantó a la de los espermistas y ovistas durante el siglo 19. La mezcla de óvulos y espermatozoides daban como resultado la progenie que era una "mezcla" ("blend") de las características de los padres. Las células sexuales se conocían colectivamente como gametos. De acuerdo con la teoría de la mezcla, cuando un animal de color negro se cruzaba con uno blanco la progenie debía ser gris y, a menudo, este no era el resultado. La teoría de la mezcla obviaba, entre otras, explicar el salto de generación de algunas características.Charles Darwin en su teoría de la evolución, se vio forzado a reconocer que la mezcla no era un factor (o al menos no el factor principal) y sugirió que la ciencia, en la mitad de los 1800s, no tenía la respuesta correcta al problema. La respuesta vino de un contemporáneo, Gregor Mendel, si bien Darwin nunca conoció el trabajo de Mendel.Conceptos básicos de la genéticaResulta útil recordar algunos conceptos previos para comprender los experimentos de Mendel, aunque este monje no haya tenido conocimiento de los genes o los cromosomas... Meiosis: división celular que origina 4 células con la mitad de la dotación cromosómica de la célula orginal (haploides). Los cromosomas homólogos se separan y cada célula (gameta) recibe uno de los homólogos del par.Carácter: característica observable y transmitida por los genes, ejemplo: color de las floresFenotipo : propiedades observables del genotipo y en el cual contribuye el medio ambiente.Cromosomas Homólogos: cromosomas que se aparean durante la meiosis. Poseen igual longitud, posición del centrómero y comparten los mismos genes. Excepción : cromosomas X e Y que no comparten las características anteriores pero sí se consideran homólogos por aparearse en la meiosis.Gen (del griego genos= nacimiento) son segmentos específicos de ADN (cromosoma) responsable de un determinado carácter; son la unidad funcional de la herencia. El botánico danés Wilhelm Johannsen (1857 - 1927) acuño este nombre, en 1909, para nombrar a los elemente de Mendel (también acuñó "fenotipo", "genotipo" y "selección").Alelo: Formas alternativas de un gen en un mismo locus. Por ejemplo 2 posibles alelos en el locus v de la cebada son v y V. El término de alelo ó alelomorfo fue acuñado por William Bateson; literalmente significa "forma alternativa".Locus: es el lugar específico de un gen en un cromosoma.Homocigoto: organismo que tiene dos copias o alelos iguales de un gen en los dos homólogos, también llamado raza pura.Heterocigoto: cuando los dos alelos son diferentes, en este caso el alelo dominante es el que se expresa.MENDELMendel razonó que un organismo apto para los experimentos genéticos debería tener :una serie de características diferentes, fácilmente estudiables y con dos o tres fenotipos diferentes.la planta debía autofertilizarse y tener una estructura floral que limite los contactos accidentales, de crecimiento rápido y con gran número de descendientes.Los descendientes de las plantas autofertilizadas debían ser fértiles.El organismo experimental de Mendel fue la arveja común (Pisum sativum, familia Leguminosae), que tiene una flor que normalmente se autopoliniza. La parte masculina de la flor se llama antera, produce el polen, que contiene los gametos masculinos. La parte femenina de la flor es el Gineceo, formado por estigma, estilo, y el ovario. El óvulo (gameto femenino) es producido en el ovario. El proceso de polinización (la transferencia de polen de la antera al estigma) ocurre, en el caso de la arveja, antes de la apertura de la flor. Del grano de polen crece un tubo (tubo polínico) que permite al núcleo viajar a través del estigma y el estilo, y eventualmente llegar al ovario. Las paredes del ovario formarán las futuras vainas (fruto: legumbre) y los óvulos fecundados las semillas. Ver ciclo animado de plantas.Muchas flores permiten la polinización cruzada, lo cual puede dificultar los estudios si se desconoce las características de la planta masculina. Dado que las flores de las arvejas el estigma y las anteras están completamente encerrados y, a diferencia de la mayoría de las flores no se abren hasta ser fecundadas, es decir luego de la autopolinización, la genética de los progenitores puede ser comprendida mas fácilmente. Los embriones autofecundados de las arvejas desarrollan sin dificultad.Para los entrecruzamientos Mendel abrió el pimpollo antes de la maduración y retiró las anteras con pinzas evitando la autopolinización. Luego las polinizó artificialmente, espolvoreando el estigma con polen recogido de otras plantas.Mendel probó las 34 variedades de arvejas disponibles a través de los vendedores de semillas. Mendel buscó caracteres con rasgos bien definidos y alternativos constantes, que constituyeran razas puras. Las arvejas de jardín fueron plantadas y estudiadas durante ocho años a fin de comprobar que el rasgo observado se mantenía constante a lo largo de varias generaciones. Así, Mendel aisló 7 pares de caracteres que eran razas puras: cada carácter estudiado se presentaba en dos variantes, tales como: altura de la planta (alta o baja), superficie de la semilla (lisa o rugosa), forma de la vaina (inflada o contraída), forma de la vaina y otras (ver esquema a continuación). En sus experimentos Mendel uso unas 28.000 plantas de arvejas.La contribución de Mendel fue excepcional en razón del enfoque metodológico utilizado para definir el problema, el uso de variables claramente entendibles y la aplicación de las matemática (estadística) al resultado experimental. Usando plantas de arvejas y el método estadístico, Mendel fue capaz de demostrar que los caracteres pasan de los padres a los hijos a través de la herencia de los genes.El principio de la segregación Mendel primero estudió la herencia de la forma de la semilla. Un cruzamiento relacionado a un solo carácter se denomina monohibridación. Mendel cruzó una raza pura de plantas con semillas lisas con una raza pura de otra que siempre producía semillas rugosas (60 fertilizaciones en 15 plantas). Todas las semillas resultantes resultaron lisas.Al año siguiente, Mendel plantó esas semillas y permitió que las mismas se autofecunden. Recogió 7324 semillas en total: 5474 lisas y 1850 rugosas. Para sistematizar el registro de datos, las generaciones fueron nombradas y numeradas. La generación parental se denomina como P. Los descendientes de la generación P son la generación F1 (la primera filial). La autofecundación de la generación de F1 produce la generación F2 (la segunda filial).P1:lisa X rugosaF1 :todas lisasF2 :5474 lisas y 1850 rugosasLo mismo sucedió con cada par de caracteres elegidos: cuando cepas puras de plantas con semillas amarillas se cruzan con razas puras de plantas con semillas verdes, todos los descendientes fueron plantas con semillas amarillas. Los padres del entrecruzamiento son la generación P1, y los descendientes representan la generación F1. Cuando los miembros de la generación F1 se entrecruzaron, Mendel recobro muchos descendientes amarillas, y algunos verdes. Luego del análisis estadístico de la generación F2, Mendel determinó que la relación entre plantas con semillas amarillas/verdes era 3:1. Las plantas con semillas verdes no aparecían en la primera generación F1, y se encontraban en la segunda F2 y sucesivas generaciones.Cruzamiento monohíbrido entre semillas amarillas (dominantes) y verdes (recesivo). Vea este Mendel concluyó que el carácter estudiado estaba gobernado por factores discretos (separables) y que el rasgo del carácter que aparece en la F1 es el dominante. Los factores se heredaban a pares, teniendo cada generación un par de los mismos. Actualmente nos referimos a esos factores como alelos. El hecho de que los caracteres se hereden de a pares permiten explicar el fenómeno observado del "salto" de una generación.Los caracteres dominantes fueron definidos por Mendel como aquellos que aparecen en la primera generación( F1) en los entrecruzamientos entre dos especies puras. Las letras mayúsculas se usan generalmente como notación para los caracteres dominantesLos caracteres recesivos son los que "saltan" una generación, y se observan únicamente cuando el carácter dominante esta ausente. Las letras minúsculas se usan generalmente como notación para los caracteres recesivos.Las plantas de Mendel exhibían dominancia completa, en las cuales las expresiones fenotípicas de los alelos eran dominantes o recesivas, sin "caracteres intermedios".La Meiosis, un proceso desconocido en los días de Mendel, explica como se heredan los caracteres:Esquema de la meiosis y formación de los gametos. Imagen modificada deSumario de los resultados de MendelLos descendientes F1 muestran solo uno de los caracteres de los padres, y siempre el mismo carácter.El carácter que no se observa en F1 reaparece en F2 en aproximadamente un 25% de los descendientes.El carácter no cambia cuando pasa a la descendencia: no se mezclan en ningún descendiente y se comportan como unidades separadas.Los cruzamientos recíprocos demostraron que cada progenitor contribuye de manera igual a la descendencia.El término fenotipo se refiere al conjunto de caracteres que se expresan o sea a la apariencia externa, mientras que el término genotipo se refiere a la totalidad genética del individuo .Machos y hembras contribuyen equitativamente a la formación del material genético de la descendencia: por lo tanto el numero de factores que determinan un carácter es probablemente dos (la solución mas simple).El Principio de la Segregación o Primera Ley de Mendel, propone la separación de los factores apareados durante la formación de los gametos, donde cada gameto recibe uno u otro factor durante su formación. Los organismos portan dos factores (alelos) por cada carácter. Estos factores se separan durante la formación de los gametos.Una versión en hipertexto (en Alemán o Ingles) del trabajo original de Mendel en 1865 se consigue siguiendo este enlace.Consecuencias de la segregaciónAlelos: se sabe ahora que cualquier gen presenta dos formas diferentes o alelos.Homo- y Heterocigosis: determinada por la combinación de los dos alelos de un gen.Fenotipo: expresión de las características genéticas o genotipo.Cuadro de PUNNETEs un mecanismo muy útil a la hora de considerar las posibles combinaciones de gametos. Por ejemplo, en la F1 todas las plantas del cruzamiento monohíbrido entre plantas altas y bajas dieron altas. El cuadro de Punnett permite calcular el resultado de la F2:Cruzamiento de pruebaPara probar la hipótesis de que los alelos están en pares y se separan en la formación de gametas se llevó a cabo un experimento adicional: se cruzó la F1 (semillas lisas) con la raza pura paterna de semillas rugosas (padre homocigota recesivo) a lo que se denominó
CRUZAMIENTO DE PRUEBA.
En un cruzamiento de prueba se cruzan un genotipo desconocido que muestra el carácter dominante con el padre homocigota recesivo. Lo que se pretende demostrar es si el genotipo desconocido es homocigota dominante o heterocigota para ese carácter. Si se producen dos fenotipos distintos quiere decir que el progenitor desconocido era heterocigota para ese carácter. Si por el contrario aparece un solo fenotipo es homocigoto.Cruzamiento dihíbridoMendel entendió que era necesario realizar su experimento en una situación más compleja y realizó experimentos siguiendo dos caracteres de las semillas: forma y color. Un entrecruzamiento concerniente a dos caracteres se conoce como cruzamiento dihíbrido en oposición al cruzamiento de una sola característica o, monohíbrido.La generación F2 resultante no muestra la característica relación fenotípica 3:1 dominante: recesivo. Los dos caracteres, si consideramos que se heredan independientemente, "calzan" dentro del principio de la segregación. En vez de los 4 posibles genotipos de un monohíbrido, el cruzamiento dihíbrido tiene 16 posibles genotipos.Cruzamientos con dos caracteresLas semillas lisas (S) son dominantes respecto a la semillas rugosos (s).El color amarillo (Y) es dominante sobre el verde (y).Una vez más, la meiosis nos ayuda a entender el comportamiento de los alelos.Métodos, Resultados y ConclusionesEl gráfico superior es de Genetics pages en McGill UniversityMendel partió de razas puras que tenían plantas con semillas lisas y amarillas, y las cruzó con razas puras de plantas con semillas verdes y arrugadas. Todas las semillas de la generación F1 tenían semillas lisas y amarillas. Las plantas de la generación F2 se obtuvieron por autofertilización, y produjeron cuatro fenotipos:315 lisas y amarillas108 lisas verdes101 arrugadas amarillas32 arrugadas verdesMendel analizó cada carácter por separado como si fuera que el otro carácter no estuviera presente. la relación 3:1 se veía separadamente y estaba de acuerdo con el Principio de Segregación. La segregación de los alelos S y s debían haber ocurrido independientemente de la separación de los alelos Y e y .La probabilidad de que un gameto tenga Y es 1/2; la probabilidad de cualquier gameto de tener S es 1/2. La probabilidad de que un gameto contenga ambos Y y S se calcula por el producto de las probabilidades individuales (o 1/2 X 1/2 = 1/4).La probabilidad de que dos gametos formen cualquier mezcla de estos alelos en su genotipo 1/4 X 1/4 (recuerde el producto de las probabilidades individuales).Por lo tanto, existen 16 posibilidades y, el tablero de Punnett tiene 16 casillas. Dado que hay mas posibilidades de combinaciones que producen el fenotipo liso y amarillo (SSYY, SsYy, SsYY, y SSYy), este fenotipo es mas común en la F2.De los resultados de su segundo experimento, Mendel formuló el Principio de la distribución independiente esto es, cuando se forman los gametos, los alelos de un gen para una característica dada se separan (segregan) independientemente de un alelo para otra característica. Si los caracteres se separan independientemente uno de otros durante la formación de los gametos, puede entenderse el resultado de un entrecruzamiento dihíbrido.Desde los tiempo de Mendel, los científicos descubrieron el cromosoma y el ADN, y actualmente se interpreta el principio de la distribución independiente como alelos de genes en diferentes cromosomas que se heredan independientemente durante la formación de los gametos. Esto no era del conocimiento de Mendel.MutaciónDe Vries en 1902 trabajando sobre la "hierba del asno" describió en ella fenómenos de herencia mendeliana, sin embargo de tanto en tanto aparecía una característica que no estaba ni en los padres ni en los antecesores de las plantas, dedujo de ello que estas característica surgían por un cambio el factor que determinaba el carácter (gen) y que este cambio se transmitía a la progenie como cualquier otro carácter hereditario. A este cambio lo denominó mutación y a los organismos que la mostraban mutantes, los alelos salvajes son los que están presentes en la mayoría de los individuos y dan el fenotipo esperado.Ni las leyes de Mendel ni el concepto de mutación fueron conocidos por Darwin, pero resulta claro que la combinación de características de los padres da resultados sobre los cuales puede actuar el proceso evolutivo y que las mutaciones (si bien raras) son una fuente constante de variaciones que posibilitan la evolución.Los diversos alelos existen porque cualquier gen está sometido a mutaciones, que ocurren cuando un gen cambia a una nueva forma estable y hereditaria. Las mutaciones son procesos aleatorios. Los alelos mutantes y salvajes residen en el mismo locus y se heredan de acuerdo a la genética mendeliana.Las Leyes de Mendel.-Primera Ley de Mendel o Principio de Ley de la Dominancia.- Esta Ley menciona que para cada característica hereditaria existen genes dominantes y recesivos. Sin importar cual padre contribuye con el carácter dominante el híbrido o heterocigoto siempre tendrá fenotipo dominante.Segunda Ley de Mendel o Principio de la Segregación de Caracteres.Un carácter hereditario se transmite como una unidad que no se combina, se diluye o se pierde al pasar de una generación a otra, sólo se segrega o se separa.Tercera Ley de Mendel o de Distribución Independiente. Anuncia que un par de alelos se distribuye en forma independiente de otro par de alelos. Los caracteres se heredan de manera independiente unos de otros.Material hereditarioEl material hereditario es un componente de las células que da las características a éstas, además de darles una actividad específica. En las células eucariontes se ubica dentro del núcleo celular. Recibe este nombre a causa de que al dividirse una célula en dos, este material se duplica y cada célula hija "hereda" una copia de él. También se le conoce como material genético.Descubrimientos a través de la historiaA mediados del siglo XIX, Gregorio Mendel determinó que el material hereditario funciona como partículas que conservan su identidad en el paso de una célula a otra al interior de los gametos. A estas "partículas" las llamó factores hereditarios.Al comenzar el siglo XX, los estudios de Mendel estaban siendo reconocidos. Había argumentos a favor de la teoría de que los factores hereditarios se encuentran dentro de una partícula llamada cromosoma. Estos pensamientos estaban favorecidos por la semejanza entre las teorías de Mendel y el comportamiento que se estaba observando microscópicamente en los cromosomas.Las investigaciones continuaron y luego, llevaron a concluir que los factores hereditarios, ahora llamados genes, estaban hechos de ADN. Una prueba de lo anterior sería que en todas las células de la misma especie, la cantidad de ADN es constante. En cambio en los gametos existe la mitad de este elemento. Ya en la década de 1960 se conocía modelos de la estructura de ésta molécula, como también se sabía como se duplicaba y como determinaba la función de una célula. Se logró establecer que la secuencia de nucleótidos presentes en el ADN determina a la secuencia de aminoácidos en las proteínas, llegando a conocer el papel que desempeña el nucleótido ARN. Para que los ribosomas logren sintetizar las proteínas, el ADN se copia mediante la transcripción, en una molécula de ARN, también llamada ARN mensajero. Este sale del núcleo y llega a los ribosomas, que a través de la traducción, leen el patrón del ARN y efectúan la síntesis de proteínas.
MUTACION DE LOS GENES
Las mutaciones fueron descritas por primera vez en 1901 por uno de los redescubridores de Mendel, el botánico holandés Hugo De Vries.Si esto se produce en la secuencia de nucleótidos que codifica un polipéptido particular, éste puede presentar un aminoácido cambiado en la cadena polipeptídica. Esta modificación puede alterar seriamente las propiedades de la proteína resultante. Cuando se produce una mutación durante la formación de los gametos, ésta se transmitirá a las siguientes generaciones. La mayoría de las mutaciones genéticas son perjudiciales para el organismo que las porta. Una modificación aleatoria es más fácil que deteriore y que no mejore la función de un sistema complejo como el de una proteína. Por esta razón, en cualquier momento, el número de sujetos que portan un gen mutante determinado se debe a dos fuerzas opuestas: la tendencia a aumentar debido a la propagación de individuos mutantes nuevos en una población, y la tendencia a disminuir debido a que los individuos mutantes no sobreviven o se reproducen menos que sus semejantes. Varias actuaciones humanas recientes, como la exposición a los rayos X con fines médicos, los materiales radiactivos y las mutaciones producidas por compuestos químicos, son responsables de su aumento.Por lo general, las mutaciones son recesivas, sus efectos perjudiciales no se expresan a menos que dos de ellos coincidan para dar lugar a una situación homocigótica. Esto es más probable en la procreación consanguínea, en el apareamiento de organismos muy relacionados que pueden haber heredado el mismo gen mutante recesivo de un antecesor común. Por esta razón, las enfermedades hereditarias son más frecuentes entre los niños cuyos padres son primos que en el resto de la población.
Mutaciones génicas.
Son las verdaderas mutaciones, porque se produce un cambio en la estructura del ADN. A pesar de todos los sistemas destinados a prevenir y corregir los posibles errores, estos pueden serlas mutaciones puntuales, que son el cambio de una de las bases de un par en el ADN, un cambio en una base nitrogenada puede alterar la estructura completa de la proteína.Tirosina arginina metioninaAUG GCU UACTirosina glutamina metioninaAUG GUU UAC
De vez en cuando se produce alguno en la réplica, bien por colocarse una Citosina (C) en lugar de una Timina (T), o una Adenina (A) en lugar de una Guanina (G); o bien porque el mecanismo de replicación se salta algunas bases y aparece una "mella" en la copia. O se unen dos bases de Timina, formando un dímero. Aunque se trate de un cambio de un nucleótido por otro, supondrá una alteración en la secuencia de un gen, que se traduce posteriormente en una modificación de la secuencia de aminoácidos de una proteína. Al transcribirse la mutación, al menos un triplete del ARNm , se encuentra modificado y su traducción da lugar a que se incorpore un aminoácido distinto del normal en la cadena polipeptídica. Es un cambio que aunque la mayoría de las veces va a ser perjudicial, en contadas ocasiones puede provocar que mejore un gen y gracias a esta característica se sintetice una proteína distinta , que tenga propiedades distintas o participe en la formación de estructuras más eficaces. En estos casos raros, pero esenciales para la evolución de las especies , los individuos portadores de la mutación poseen ventajas adaptativas respecto a sus congéneres , por lo que el gen mutado es posible que con el tiempo, y gracias a la selección natural, sustituya al gen original en la mayoría de los individuos que componen la población
Mutaciones cromosómicas
La sustitución de un nucleótido por otro no es el único tipo posible de mutación. Algunas veces se puede ganar o perder por completo un nucleótido. Además, es posible que se produzcan modificaciones más obvias o graves, o que se altere la propia forma y el número de los cromosomas. Una parte del cromosoma se puede separar, invertir y después unirse de nuevo al cromosoma en el mismo lugar. A esto se le llama inversión. Si el fragmento separado se une a un cromosoma distinto, o a un fragmento diferente del cromosoma original, el fenómeno se denomina translocación. Algunas veces se pierde un fragmento de un cromosoma que forma parte de una pareja de cromosomas homólogos, y este fragmento es adquirido por el otro. Entonces, se dice que uno presenta una deleción o deficiencia (dependiendo si el fragmento que se pierde es intersticial o terminal, respectivamente) y el otro una duplicación. Por lo general, las deficiencias o deleciones son letales en la condición homocigótica, y con frecuencia las duplicaciones también lo son. Las inversiones y las translocaciones suelen ser más viables, aunque pueden asociarse con mutaciones en los genes cerca de los puntos donde los cromosomas se han roto. Es probable que la mayoría de estos reordenamientos cromosómicos sean la consecuencia de errores en el proceso de sobrecruzamiento.
Errores de dusyución
Este tipo de mutaciones afectan a la dotación cromosómica de un individuo, es decir, los individuos que las presentan tienen en sus células un número distinto de cromosomas al que es propio de su especie. No son mutaciones propiamente dichas, porque no hay cambio de material genético, sino una aberración, la cual suele ser el resultado de una separación anormal de los cromosomas durante la meiosis, con lo que podemos encontrarnos individuos triploides (3n), tetraploides (4n), etc. Cuando en la meiosis fracasa la separación de una pareja de cromosomas homólogos. Esto puede originar gametos —y, por lo tanto, cigotos— con cromosomas de más, y otros donde faltan uno o más cromosomas. En el hombre, existen varios síndromes provocados por la no separación de una pareja de cromosoma homólogos durante la meiosis, con lo cual permanecen unidos y se desplazan juntos a un mismo gameto provocando lo que se denomina trisomía, es decir un individuo con un cromosoma triplicado. Los individuos con un cromosoma de más se denominan trisómicos, y aquellos en los que falta uno, monosómicos. Ambas situaciones tienden a producir incapacidades graves. Por ejemplo, las personas con síndrome de Down son trisómicas, con tres copias del cromosoma 21.En la meiosis fracasa a veces la separación de un grupo completo de cromosomas; es decir, se origina un gameto con el doble del número normal de cromosomas. Si dicho gameto se une con otro que contiene el número normal de cromosomas, el descendiente tendrá tres grupos de cromosomas homólogos en lugar de los dos habituales. Si se unen dos gametos con el doble del número normal de cromosomas, el descendiente estará dotado de cuatro grupos homólogos. Los organismos con grupos adicionales de cromosomas reciben el nombre de poliploides. La poliploidía es el único proceso conocido por el cual pueden surgir especies nuevas en una generación única. Se han observado poliploides viables y fértiles casi exclusivamente en organismos hermafroditas, como la mayoría de las plantas con flores y algunos invertebrados Por lo general, las plantas poliploides son mayores y más robustas que sus antecesoras diploides Estos poliploides así formados son genéticamente muy interesantes en las plantas cultivadas, y hoy en día la mayoría de variedades gigantes de fresones, tomates, trigo, ... que existen en el mercado, tienen este origen.. Algunas veces se originan fetos poliploides en la raza humana, pero fallecen en una fase precoz del desarrollo fetal y se produce un aborto.En los siguientes esquemas, tenemos las trisomías más frecuentes tanto en los autosomas, como en los cromosomas sexuales.
Suscribirse a:
Entradas (Atom)
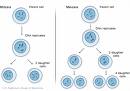
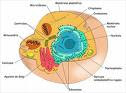
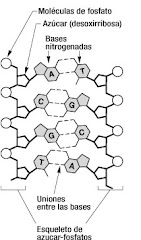

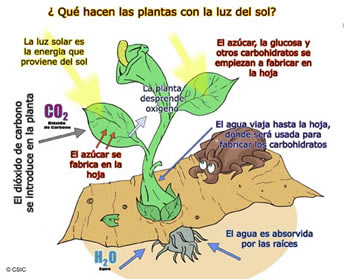
{kind=link}